
C точки зрения медицинской генетики эссенциальная гипертензия, или гипертоническая болезнь (ГБ), представляет собой мультифакторное полигенное заболевание, развитие которого определяется сложным характером взаимодействия множества генов с разнообразными факторами окружающей среды [1, 2]. На протяжении последнего десятилетия, благодаря успешной расшифровке генома человека и разработке высокоразрешающих и производительных методов генотипирования полиморфизмов в молекуле ДНК, достигнуты значительные успехи в выяснении генетических причин ГБ, определены многочисленные гены предрасположенности к заболеванию и открыты ранее неизвестные стороны его патогенеза [1—4]. Установлено, что генетическую детерминацию ГБ составляет большое число полиморфных генов со слабым или умеренным фенотипическим эффектом, а их разнообразные комбинации у разных людей лежат в основе существенных различий по патогенезу (генетическая гетерогенность) и клинической картине (клинический полиморфизм) болезни не только на межрасовом или межэтническом уровнях, но даже в пределах одной популяции [1, 2, 5, 6].
Хорошо известно, что межиндивидуальные и межпопуляционные различия молекулярно-генетических механизмов артериальных гипертензий (АГ), как, впрочем, и других мультифакторных болезней, а также специфичность влияний факторов внешней среды на отдельно взятые популяции, модифицирующая фенотипические эффекты генов (взаимодействия генотип—среда) [7—9], существенно затрудняют выявление новых и инвентаризацию известных генов-кандидатов подверженности ГБ. В этой связи возникает необходимость тестирования генетических ассоциаций маркеров с ГБ с учетом известных средовых факторов риска (ФР) развития заболевания, что позволяет не только охарактеризовать особенности молекулярных механизмов его развития в конкретной популяции, но и определить на их основе спектр средовых ФР, провоцирующих возникновение данной патологии в регионе.
В первую десятку наиболее часто исследуемых полиморфных генов в отношении риска развития АГ в различных популяциях мира входит ген α-аддуцина (ADD1) [10]. Ген α-субъединицы аддуцина картирован на коротком плече 4-й хромосомы в сегменте 16.3, который кодирует белок молекулярной массой 200 кБа [11]. Наиболее хорошо изученным и функционально важным полиморфным участком гена ADD1является мутация в 10-м экзоне, характеризующаяся аминокислотной заменой глицина Gly (G) на триптофан Trp (W) в 460-м положении полипептидной цепи белка (Gly460Trp, G460W) [10]. В некоторых европеоидных и монголоидных популяциях выявлена связь аллельного варианта 460W с развитием ГБ, повышением уровня артериального давления (АД) [12—15] и его снижением на фоне ограничения употребления поваренной соли [10] и терапии тиазидными диуретиками [16]. Однако в других популяциях мира ассоциацию полиморфизма G460W гена ADD1с риском развития ГБ и уровнем АД выявить не удалось [17—20]; это служит демонстрацией межпопуляционных различий во взаимосвязи гена α-аддуцина и риска развития заболевания.
Цель настоящей работы состояла в исследовании связи полиморфизма G460W гена ADD1 с риском развития ГБ, а также роли средовых ФР в реализации предрасположенности к заболеванию у носителей вариантного генотипа данного гена.
Материал и методы
Материалом для исследования послужила популяционная выборка неродственных индивидуумов русской национальности, проживающих в Центральной России — 412 коренных жителей Курской области. Были обследованы 205 больных ГБ, которые находились на стационарном лечении в кардиологических отделениях Курской областной клинической больницы и городской больницы скорой медицинской помощи Курска за период с 2003 по 2004 г. Контрольная группа включала 207 практически здоровых добровольцев и была сформирована при профилактических осмотрах на протяжении того же периода времени. Большинство больных длительно страдали ГБ и получали антигипертензивную терапию. У первичных больных диагноз ГБ устанавливали согласно рекомендациям ВОЗ после исключения симптоматических АГ с помощью клинических, лабораторных и клинико-инструментальных методов. Критерием включения здоровых индивидуумов в контрольную группу были систолическое АД — 130 мм рт.ст., диастолическое АД — 85 мм рт.ст. при двух повторных измерениях, отсутствие выраженных хронических заболеваний. Группы здоровых и больных ГБ были сопоставимы по полу и возрасту (р>0,05). Средний возраст больных ГБ (67 мужчин, 168 женщин) составил 47,8±9,6 года, здоровых индивидуумов (75 мужчин, 132 женщин) — 45,9±11,2 года. Согласно оценке тяжести АГ больные распределялись следующим образом: у 81,4% имелась АГ 3-й степени, у 12,0% — АГ 2-й степени и у 6,6% больных — АГ 1-й степени. У 58,7% больных выявлена лабильная АГ, у 41,3% — стабильная форма. У 22,3% больных ГБ характеризовалась кризовым течением.
Анкетирование проводили по специально разработанной анкете при непосредственном общении с пациентами. Кроме основных социально-демографических параметров опросник включал сведения о наличии у обследуемых средовых ФР развития ГБ, таких как вредные привычки (курение, злоупотребление алкоголем), характер питания («солевой аппетит», уровень употребления свежих овощей/фруктов), наличие хронических стрессовых ситуаций и уровень физической активности. Относительно статуса курения больные подразделялись на 2 группы: курящие (курил(а) и продолжает курить) и некурящие (никогда не курил(а)). Наличие в анамнезе хронических стрессовых ситуаций выявляли на основании ответов на вопрос: «была ли связана Ваша жизнь с постоянными стрессами?» (ответы: «да» и «нет»). Уровень физической активности оценивали по 3 категориям: 1) характер трудовой деятельности: а) работа не связана с физической деятельностью/сидячая работа — 0 баллов; б) работа связана с умеренной физической нагрузкой — 1 балл; в) работа связана с высокой физической нагрузкой — 2 балла; 2) занятия спортом в свободное время: а) спортом не занимаюсь — 0 баллов; б) спортом занимаюсь эпизодически/нерегулярно — 1 балл; в) систематическое занятие спортом — 2 балла; 3) средняя продолжительность времени, затрачиваемого на пешеходную прогулку, в течение дня: а) менее 30 мин в день — 0 баллов; б) от 30 мин до 1 ч в день — 1 балл; в) 1—2 ч в день — 2 балла; г) более 2 ч в день — 3 балла. Оценка у респондентов по всем категориям ответов 2 балла или менее расценивалась как низкая физическая активность, оценка более 2 баллов — как умеренная/ высокая физическая активность. Градация «употребление алкоголя» (крепких алкогольных напитков) по частоте его употребления ранжировалась на следующие категории: 1) реже 1 раза в месяц или не употребляю; 2) 1—2 раза в месяц; 3) 1—2 раза в неделю; 4) 3—4 раза в неделю; 5) ежедневное употребление алкоголя. Респондентов, выбравших 4-ю или 5-ю категории ответов, относили к лицам с высоким уровнем употребления алкоголя; респондентов, выбравших с 1-й по 3-ю категории ответов, — к лицам с умеренным/низким уровнем употребления алкоголя. «Солевой аппетит» у респондентов оценивали посредством вопроса о предпочтении к употреблению соленой пищи по 3 категориям ответов: 1) «предпочитаю слегка несоленую пищу»; 2) «предпочитаю умеренно соленую пищу»; 3) «предпочитаю пересоленную пищу». Затем из респондентов были сформированы 2 группы: 1- я группа — лица с высоким «солевым аппетитом» (3-я категория ответов); 2-я группа — лица с умеренным или низким «солевым аппетитом» (1-я и 2-я категории ответов). Уровень употребления свежих овощей и фруктов «как часто Вы употребляете свежие овощи и фрукты?» (вопрос был выбран из пищевого опросника, разработанного National Cancer Institute [21]) оценивали по следующим категориям: 1) не употребляю; 2) 1—3 раза в месяц; 3) 1— 2 раза в неделю; 4) 3—4 раза в неделю; 5) 5—6 раз в неделю; 6) 1—2 раза в день; 7) 3 раза в день; 8) 4 раза в день; 9) 5 раз в день и более. Затем формировали две группы: 1-я группа — лица с высоким уровнем употребления свежих овощей и фруктов (2 раза в день и более); 2-я группа — лица с умеренным или низким уровнем употребления свежих овощей и фруктов (1 раз в день и реже).
У всех обследуемых брали образцы венозной крови в количестве 5—10 мл в пластиковые пробирки с 0,5М ЭДТА, после чего образцы замораживали и хранили при температуре —20 °С. Геномную ДНК выделяли из размороженной крови стандартным методом фенольно-хлороформной экстракции с последующей преципитацией этанолом. Генотипирование полиморфизма G460W (rs4961) гена ADD1 осуществляли методами полимеразной цепной реакции (ПЦР) и рестрикционного анализа с использованием праймеров, описанных в работе A.S. Morrison и соавт. [22]. ПЦР проводили в 12 мкл реакционной смеси на многоканальном термоциклере Терцик (НПО ДНК-Технология, Москва). Смесь для амплификации включала 67 мМ Трис-HCl pH 8,8, 16,6 мМ сульфата аммония, 6,7 мкМ ЭДТА, 2,5 мМ MgCl2, 1 мМ α-меркаптоэтанола, 1 мМ каждого dNTPs (dATP, dCTP, dTTP и dGTP), 15 нМ каждого из праймеров и 1U Taq-полимеразы. Продукт ПЦР размером 147 пар нуклеотидов подвергали гидролизу эндонуклеазой AspS9I (Сибэнзим, Новосибирск), после чего фрагменты ДНК фракционировали в 2% агарозном геле, окрашенном бромистым этидиумом, и документировали на трансиллюминаторе в проходящем ультрафиолетовом свете с применением компьютерной видеосъемки на приборе GDS-8000 (UVP, США). Образцы электрофоретического разделения продуктов ПЦР-рестрикции полиморфизма G460W гена ADD1 представлены на рис. 1.
Рисунок. Образец электрофоретического разделения фракментов ДНК при генотипировании полиморфизма 460GW гена ADD1.
Слева обозначены размеры фрагментов ДНК (пн — пар нуклеотидов) после рестрикции продукта ПЦР эндонуклеазой AspS9I. Генотип 460W/W визуализируется на геле в виде одной верхней полосы размером 147 пн, генотип 460G/G — в виде одной нижней полосы размером 122 пн, гетерозиготный генотип 460G/W — в виде двух полос размерами 147 и 122 пн (ДНК-фрагмент размером 25 пн не визуализируется).
Статистическую обработку полученных данных проводили с использованием программных пакетов Statistica 6.0 (StatSoft, США) и Excel 2002 (Microsoft, США). Для оценки ассоциаций аллелей и генотипов G460W гена ADD1с предрасположенностью к ГБ использовали критерий χ2и отношение шансов (ОШ) [23] — показатель, отражающий, во сколько раз вероятность оказаться в группе «случай» (больные ГБ) отличается от вероятности оказаться в группе «контроль» (здоровые) для носителя изучаемого генотипа. При ОШ>1 судили об отсутствии ассоциации; ОШ=1 рассматривалось как наличие положительной ассоциации ДНК-маркера (ФР) и ГБ; ОШ<1 свидетельствовало об отрицательной ассоциации ДНК-маркера (фактор устойчивости) с ГБ. Границы 95% доверительного интервала (ДИ) для ОШ вычисляли методом B. Woolf. Анализ взаимодействий генотип—среда проводили путем сопоставления величин ОШ, рассчитанных для генотипов в группах, которые были сформированы по критерию наличия или отсутствия средового ФР. Различия считали статистически значимыми при p≤0,05.
Результаты
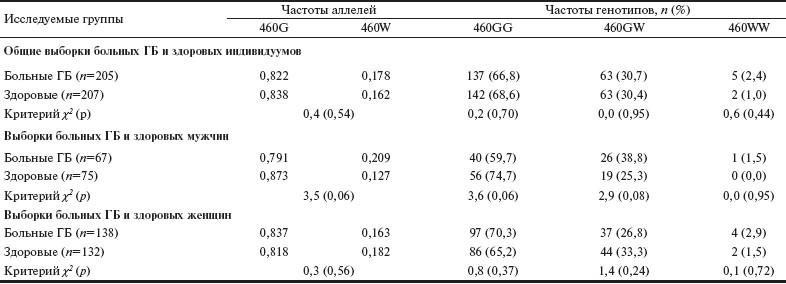
Частоты генотипов полиморфизма G460W гена ADD1 находились в соответствии с равновесием Харди— Вайнберга как в контрольной группе, так и в группе больных ГБ (p>0,05). В табл. 1 представлены частоты аллелей и генотипов полиморфизма G460W гена ADD1 в исследованных группах. Статистически значимых различий по частотам аллелей полиморфизма G460W гена ADD1 между группами здоровых и больных ГБ не выявлено. Частота аллеля 460W в исследуемой русской популяции не отличалась от таковой в других европеоидных популяциях [5, 24]. Стратифицированный сравнительный анализ частот аллелей и генотипов в зависимости от пола также не выявил статистически значимых различий в их распределении между группами здоровых и больных ГБ мужчин и женщин. Однако среди мужчин наблюдалась отчетливая тенденция ассоциации аллеля 460W (ОШ 1,82 при 95% ДИ от 0,96 до 3,44; p=0,062) и вариантных генотипов 460GW+WW (ОШ 1,99 при 95% ДИ от 0,97до 4,06; p=0,057) гена ADD1 с повышенным риском развития ГБ, но различия между группами не достигали уровня статистической значимости.
Таблица 1. Частоты аллелей и генотипов полиморфизма G460W гена ADD1 среди больных ГБ и здоровых индивидуумов.
Примечание. Здесь и в табл. 2: ГБ — гипертоническая болезнь.
В дальнейшем мы проанализировали взаимодействия вариантного генотипа 460GW с такими средовыми ФР, как курение, злоупотребление алкоголем, повышенный «солевой аппетит», низкий уровень употребления свежих овощей и фруктов, наличие хронических стрессов и низкий уровень физической активности. Среди больных ГБ было существенно меньше (p=0,002) курильщиков (n=30; 17,2%), чем в контрольной группе (n=50; 32,1%). Число лиц, злоупотребляющих алкоголем (употребляющих алкоголь чаще 3 раз в неделю), также было значительно меньше (p=0,0001) среди больных ГБ (n=28; 16,4%), чем среди здоровых индивидуумов (n=41; 36,3%). Напротив, больные ГБ (n=50; 33,8%) существенно чаще, чем здоровые (n=10; 11,0%), отмечали наличие в анамнезе хронических стрессовых ситуаций (p=0,0001). Кроме того, больные ГБ (n=107; 72,3%) значительно чаще, чем здоровые (n=46; 49,5%), отмечали низкий уровень физической активности (p=0,0003). По уровню употребления свежих овощей и фруктов, а также «солевому аппетиту» статистически значимых различий между группами не выявлено.
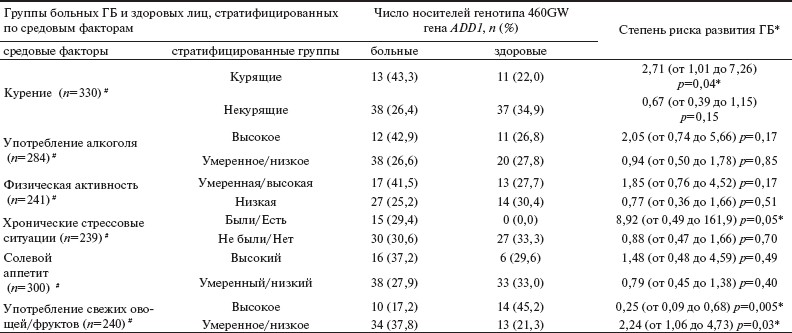
Результаты анализа взаимодействий генотип—среда представлены в табл. 2. У курильщиков — носителей генотипа 460GW гена ADD1 — был повышен риск развития ГБ (ОШ 2,71 при 95% ДИ от 1,01 до 7,26; p=0,04), тогда как у некурящих индивидуумов данный генотип не оказывал негативного влияния на риск возникновения заболевания (ОШ 0,67 при 95% ДИ от 0,39 до 1,15; p=0,15). Кроме того, у лиц, отмечавших постоянные стрессовые ситуации, наблюдалась отчетливая тенденция (p=0,056) к увеличению риска развития ГБ, но исключительно у обладателей генотипа 460GW гена ADD1. Было также установлено, что у носителей генотипа 460GW гена ADD1, не употребляющих или употребляющих в недостаточном количестве свежие овощи и фрукты, имелся наиболее высокий риск развития ГБ (ОШ 2,24 при 95% ДИ от 1,06 до 4,73; p=0,03), в то время как у лиц, регулярно употребляющих свежие овощи и фрукты, данный генотип обладал протективными свойствами в отношении риска развития заболевания (ОШ 0,25 при 95% ДИ от 0,09 до 0,68; p=0,005). Не было установлено ассоциаций генотипа 460GW гена ADD1 с риском развития ГБ в зависимости от уровня употребления алкоголя, характера «солевого аппетита» и уровня физической активности.
Таблица 2. Зависимость риска развития ГБ у носителей гетерозиготного генотипа 460GW гена ADD1 от влияния различных средовых факторов.
Примечание. #— число обследованных лиц, проанкетированных по данному средовому фактору; * — данные представлены в виде ОШ (95% ДИ).
Обсуждение
Аддуцин представляет собой гетеродимерный белок цитоскелета клеточных мембран, который посред- цством связывания с Ca2+-кальмодулином обеспечивает формирование комплексов спектрин—актин и участвует в трансмембранной передаче сигналов, являясь субстратом для протеинкиназ С и А [25, 26]. Аддуцин вовлечен в регуляцию ферментативной активности Na+/K+-ATфазы клеток почечных канальцев, и таким образом участвуя в процессах транспорта натрия через клеточную мембрану [27]. Согласно данным некоторых исследований, ген ADD1 экспрессируется во многих органах и тканях человека, включая клетки мозга и почек, эндотелиальные и гладкие мышечные клетки, кардиомиоциты и эритроциты. Известно, что функциональный эффект аминокислотной замены Gly460Trp гена ADD1проявляется перераспределением межбелковых взаимодействий в структуре цитоскелета, нарушением полимеризации актина, увеличивая активность Na+/K+-ATфазы, способствуя тем самым усилению канальциевой реабсорбции натрия в почках и формированию гипоренинной формы АГ [27, 28]. Выдвинутые предположения о возможной вовлеченности α-аддуцина в молекулярные механизмы первичной АГ послужили основанием для изучения влияния полиморфизма G460W гена ADD1 на риск возникновения ГБ в различных популяциях мира. Однако многочисленные исследования в данном направлении дали противоречивые результаты, свидетельствующие о том, что связь полиморфизма G460W гена ADD1 с риском возникновения ГБ может носить популяционноспецифический характер и зависеть от модифицирующих влияний определенных средовых факторов.
В рамках настоящего исследования впервые в русской популяции было изучено влияние полиморфизма G460W гена ADD1 на риск развития ГБ. Хотя мы выявили взаимосвязи данного полиморфизма с развитием ГБ при сопоставлении групп больных и здоровых индивидуумов, последующий стратифицированный анализ показал, что в исследованной русской популяции генотип 460GW является значимым фактором повышенного риска возникновения заболевания у курильщиков и лиц, употребляющих недостаточное количество свежих овощей и фруктов. Было также показано, что у носителей генотипа 460GW на фоне хронического стресса повышен риск развития ГБ.
Полученные результаты представляют дополнительные доказательства о значимой роли взаимодействий генотип—среда в реализации наследственной предрасположенности к ГБ как типичному примеру мультифакторного полигенного заболевания. Примечательно, что средовые факторы, для которых нами было установлено синергичное с генотипом 460GW гена ADD1 влияние на риск развития ГБ, представляют собой известные ФР заболевания. Так, давно не подлежит сомнению, что хронические стрессы оказывают негативное влияние на развитие ГБ [29]. Однако с учетом полученных нами результатов не совсем понятен механизм, посредством которого хронический стресс реализует свое патологическое влияние у носителей генотипа 460GW гена ADD1. С патогенетической точки зрения функциональный эффект гетерозиготного генотипа 460GW, как известно из литературы [28], связан с повышенной чувствительностью АД к приему поваренной соли и увеличением реабсорбции натрия и воды в почках, являясь основой для развития солезависимой АГ. Однако связь полиморфизма G460W гена ADD1с чувствительностью АД к приему поваренной соли не была подтверждена в отдельных исследованиях [20]. По-видимому, на фоне постоянной активации симпатико-адреналовой системы, которая физиологически интегрирована с ренин-ангиотензин-альдостероновой системой, у лиц с данным генотипом почки переключаются на режим усиленной задержки натрия и воды в организме, что и приводит к развитию АГ. Вне всякого сомнения, факт того, что мы не выявили модифицирующего влияния «солевого аппетита» на риск развития ГБ у носителей генотипа 460GW, абсолютно не исключает возможности взаимосвязи уровня потребления поваренной соли и полиморфного гена α-аддуцина в детерминации уровня АД и ГБ. Конечно, для более точной оценки уровня потребления поваренной соли необходимы количественный анализ ее содержания в рационе, измерение параметров водно-солевого гомеостаза в крови и моче, а также измерение порога вкусовой чувствительности к соли, что позволит подойти к оценке «солевого аппетита» как комплексного фенотипа, имеющего патогенетическое значение для развития АГ.
Наиболее интересными и заслуживающими внимания находками были обнаруженные нами модифицирующие влияния средовых факторов прооксидантного (курение) и антиоксидантного (употребление свежих овощей и фруктов) действия на формирование риска развития ГБ у носителей гетерозиготного генотипа 460GW гена ADD1. В доступной отечественной и зарубежной литературе мы не обнаружили подобных результатов, демонстрирующих влияние курения и употребления свежих овощей и фруктов на ассоциацию полиморфизма G460W гена ADD1с развитием ГБ. Примечательно, что эффект указанных средовых факторов носил отчетливое разнонаправленное влияние на риск развития ГБ у носителей гетерозиготного генотипа 460GW. Так, у носителей генотипа 460GW на фоне курения наблюдался повышенный риск развития ГБ, тогда как у некурящих индивидуумов данный генотип был нейтрален в отношении риска возникновения заболевания. Одним из общеизвестных механизмов негативного действия компонентов табачного дыма является стимуляция симпатико-адреналовой и ренин-ангиотензин-альдостероновой систем, активация которых может опосредовать гипертензивный эффект табачного дыма через описанный выше объемзависимый механизм в эпителии почечных канальцев, где экспрессируется белок аддуцин.
Несколько иной характер ассоциации генотипа 460GW гена ADD1 с ГБ наблюдался в зависимости от уровня употребления свежих фруктов и овощей. Так, при умеренном или низком уровне употребления свежих фруктов и овощей (1 раз в день или реже) у гетерозиготных носителей полиморфизма G460W гена ADD1выявлен повышенный риск развития ГБ. На фоне высокого потребления свежих фруктов и овощей данный генотип, напротив, обладал протективными свойствами в отношении ГБ. Последняя особенность наглядно демонстрирует антигипертензивную роль употребления пищи растительного происхождения, которая проявляет свое защитное действие даже при наличии у индивидуума генотипа повышенного риска развития ГБ. По всей видимости, при недостаточном поступлении в организм природных антиоксидантов (низкий уровень потребления свежих овощей и фруктов) и на фоне постоянной экспозиции организма прооксидантами (курение) у носителей генотипа 460GW гена ADD1 создаются условия для нарушения эндогенного баланса оксидантов/антиоксидантов, усиления свободнорадикальных процессов и индукции окислительного стресса — одного из важнейших звеньев патогенеза ГБ [30].
Следует отметить, что подобные генно-средовые взаимодействия были нами недавно обнаружены и при изучении связи полиморфизма генов каталазы [31] и НАДФН-оксидазы [32] (эти гены известны как гены-кандидаты ГБ) с риском развития бронхиальной астмы, в развитии которой также немаловажную роль играют нарушения свободнорадикальных процессов и усиление окислительного стресса. В целом, обсуждая природу установленных нами генно-средовых взаимодействий, затрагивающих ген α-аддуцина и различные факторы внешней среды, следует отметить, что данные взаимодействия, по-видимому, носят неспецифический характер и представляют собой проявление неспецифического адаптивного механизма у индивидуумов с вариантным генотипом к разнообразным и сопряженным в своих действиях на организм средовым влияниям, являющимся неотъемлемыми элементами современной цивилизации.
Заключение
Полиморфизм G460W гена ADD1 может рассматриваться в качестве гена предрасположенности к ГБ в русской популяции. Причем патологическое действие данного гена проявляется исключительно при определенных условиях потенциально регулируемых факторов внешней среды. Иными словами, носительство «патологического» генотипа 460GW гена ADD1 является недостаточным для реализации наследственной предрасположенности к ГБ, а определенные средовые факторы выступают в качестве пусковых механизмов, потенцирующих развитие патологии. Биологический смысл выявленных ассоциаций гена ADD1расширяет наши представления о механизмах реализации наследственной предрасположенности не только к ГБ, но и к другим сердечно-сосудистым заболеваниям мультифакторной природы. Результаты настоящей работы обосновывают необходимость обязательного включения разнообразных средовых ФР при тестировании ассоциаций генов предрасположенности к ГБ. Полученные результаты могут иметь прямое практическое применение в клинической кардиологии, так как на молекулярногенетическом уровне обосновывают необходимость более широкого внедрения элементов превентивной антиоксидантной терапии и профилактики сердечно-сосудистой патологии, которые уже в ближайшей перспективе могут стать серьезной альтернативой современным методам антигипертензивного лечения.
Работа выполнена в рамках федеральной целевой программы «Научные и научно-педагогические кадры инновационной России» на 2009—2013 гг.



{kind=link}
{kind=link}
{kind=link}