
В условиях сниженного образования макроэргических фосфатов и перегрузки ионами Са2+ образование активных форм кислорода (АФК) способно инициировать необратимые повреждения клеточных структур — липидов, белков и ДНК, приводящие к постишемической гибели кардиомиоцитов [1]. В связи с этим разработка соединений, снижающих реперфузионные повреждения миокарда, является актуальной задачей современной экспериментальной кардиологии. В последние годы возрос интерес к изучению С-концевого фрагмента адипокина апелина (H-Arg-Pro-Arg-Leu-Ser-His-Lys-Gly-Pro-Met-Pro-Phe-OH — A12), обладающего кардипротекторной активностью. Показано, что под действием этого пептида происходят уменьшение размеров инфаркта миокарда (ИМ) у крыс in vivo и улучшение восстановления сократительной функции изолированного перфузируемого сердца крысы после ишемии [2, 3]. Однако наличие активных амино- и карбоксипептидаз в крови уменьшает период полувыведения пептида из кровотока до нескольких минут, что затрудняет применение A12 в клинике [4]. Недавно нами синтезирован структурный аналог апелина-12 H-(NαMe)Arg-Pro-Arg-Leu-Ser-His-Lys-Gly-Pro-Nle-Pro-Phe-OH (АI). Его протеолитическая устойчивость обусловлена включением в N-концевую часть пептида остатка Nα-метиларгинина, а большая стабильность — заменой метионина устойчивым к окислению кислородом норлейцином. На моделях ишемического и реперфузионного повреждения сердца показано, что AI по эффективности защиты миокарда сопоставим с природным пептидом [5, 6].
Механизмы защитного действия С-концевых фрагментов апелина изучены недостаточно. Полагают, что связываясь с рецептором APJ, они могут активировать компоненты сигнальных каскадов, включающие PI-3-Akt киназы и МАР-киназы, мишенями которых являются эндотелиальная NO-синтаза (eNOS), рибосомальная p70 S6-киназа и проапоптозный белок BAD/Bax [7, 8]. Результатом этого является ингибирование открытия митохондриальной поры, вызывающего летальные повреждения клеток, которые приводят к некрозу и апоптозу. Помимо ингибирования открытия митохондриальной поры, фрагменты апелина могут снижать реперфузионное повреждение сердца вследствие проявления ими антиоксидантного действия. Так, в культуре кардиомиоцитов при моделировании гипоксии и реоксигенации экзогенный апелин-13 уменьшал образование вторичного продукта свободнорадикального окисления липидов — малонового диальдегида (МДА) и супероксидного анион-радикала (О2), а также увеличивал экспрессию антиоксидантного фермента супероксиддисмутазы (СОД), утилизирующей О2-[9]. Меньшие повреждения мембран кардиоми-оцитов и снижение продукции МДА отмечены при защите апелином-13 поврежденного изопротеренолом миокарда у крыс in vivo [10]. Показано, что подавление пироглутаматным апелином-13 ([Pyr1]апелин-13) окислительного стресса, индуцированного пероксидом водорода (Н2О2) в изолированных кардиомиоцитах, сопровождается повышением уровня мРНК и активности каталазы [11]. Снижение гипертрофии левого желудочка (ЛЖ) у мышей под действием диеты, обогащенной [Pyr1]апелином-13, сопровождалось уменьшением концентрации липогидропероксидов в плазме и увеличением активности каталазы в миокарде [11]. Можно полагать, что уменьшение реперфузионного повреждения под действием пептидов A12 и АI связано со снижением генерирования АФК, вызванного изменением экспрессии генов СОД и каталазы или увеличением продукции NO. Участие NO в защите миокарда подтверждено уменьшением ограничения размеров ИМ пептидом А12 при ингибировании активности NOS L-NAME [3] и, вероятно, связано с антиоксидантными свойствами NO. Влияние апелинов A12 и АI на активность глутатионпероксидазы (ГП) до настоящего времени не изучено.
В связи с этим цели настоящей работы состояли в изучении влияния пептидов А12 и его структурного аналога АI на активность СОД, каталазы и ГП, а также на образование АФК и МДА в сердце крыс при реперфузии. Для исключения эффектов, обусловленных активностью аминопептидаз плазмы и нейрогуморальным влиянием, часть опытов выполнена на изолированном сердце крыс, перфузируемом модифицированным буфером Кребса-Хензелейта. Способность А12 ограничивать размеры ИМ сопоставлена с его влиянием на систему антиоксидантной защиты в сердце крыс in vivo.
Материал и методы
Получение апелина-12 и его структурного аналога. Апелин-12 (H-Arg-Pro-Arg-Leu-Ser-His-Lys-Gly-Pro-Met-Pro-Phe-OH – А12) и его структурный аналог Nα-MeArg-Pro-Arg-Leu-Ser-His-Lys-Gly-Pro-Nle-Pro-Phe-OH (AI) были получены путем автоматического твердофазного синтеза на пептидном синтезаторе фирмы «Applied Biosystems 431A» с использованием Fmoc-методологии на полимере Ванга по стандартным программам. Пептиды были очищены с помощью высокоэффективной жидкостной хроматографии до 98% чистоты, их структура была охарактеризована с помощью 1Н-ЯМР-спектроскопии и MALDI-масс-спектрометрии. В работе использованы производные аминокислот, реагенты и растворители фирм «Bachem» и «Fluka» [5].
Перфузия изолированного сердца крысы. Опыты выполнены на сердце крыс-самцов линии Вистар (290–340 г). У наркотизированных уретаном (внутрибрюшинно 1,25 мг на г массы тела) животных извлекали сердце и перфузировали ретроградно в течение 15–20 мин раствором Кребса с 11 мМ глюкозой, насыщенным карбогеном (95% О2+5% СО2), рН 7,4±0,1 при температуре 37 °С, при постоянном перфузионном давлении 60 мм рт. ст. После этого сердца перфузировали антеградно по Нийли при постоянном давлении наполнения левого предсердия 15 мм рт. ст. и среднем перфузионном давлении в аорте 60 мм рт. ст. [2]. Давление в аорте и ЛЖ регистрировали при помощи тензометрических датчиков Р 50, монитора SP 1405 и регистратора SP 2010. Показателем интенсивности сократительной функции (ИСФ) ЛЖ служило произведение частоты сокращений сердца на развиваемое давление (разность между систолическим и минимальным диастолическим давлением). Насосную функцию ЛЖ оценивали по величине минутного объема (MO – сумма коронарного потока и аортального объема).
После перфузии сердца по Нийли в течение 20 мин регистрировали показатели функции сердца и коронарных сосудов (исходное состояние). Затем произвольным образом сердца включали в одну из трех групп. В контроле осуществляли 5-минутную инфузию раствора Кребса с постоянной скоростью 4 мл/мин и подвергали сердца глобальной нормотермической (37 °С) ишемии в течение 35 мин; за ишемией следовала 5-минутная ретроградная инфузия раствора Кребса со скоростью 4 мл/мин и реперфузия по Нийли в течение 25 мин. Пептиды растворяли в растворе Кребса; растворы вводили в сердце путем инфузии со скоростью 4 мл/мин в течение 5 мин до ишемии: в группе A12 – 140 мкМ А12, в группе AI – 140 мкМ АI. При выборе концентраций приняты во внимание результаты работы [2]. В конце реперфузии или перед ишемией (исходное состояние) сердца замораживали охлажденными в жидком азоте щипцами Волленбергера для последующего определения активности Сu, Zn-CОД, каталазы, ГП и содержания МДА.
Региональная ишемия и реперфузия сердца крыс in vivo. У наркотизированных 20% уретаном (120 мг/кг массы тела внутрибрюшинно) самцов крыс линии Вистар массой 300–350 г в условиях трахеотомии осуществляли искусственную вентиляцию легких комнатным воздухом с помощью аппарата KTR-5. Яремную вену катетеризировали для окрашивания сердца 1% раствором Эванса в конце опыта, сонную артерию — для регистрации артериального давления (АД). Среднее АД и частоту сокращений сердца регистрировали в режиме on-line с помощью полиграфа Biograph-4. Запись на компьютер выполняли, используя аналого-цифровой преобразователь USB 6210 и программу LabView 7. После окончания препарирования животного следовал 30-минутный период стабилизации гемодинамических показателей (исходное состояние), затем – окклюзия передней нисходящей коронарной артерии (ПНА) в течение 40 мин; продолжительность последующей реперфузии составляла 60 мин. Для подтверждения моделирования региональной ишемии, вызванной окклюзией ПНА, использовали показания электрокардиографии в отведении, соответствующем II.
Пептид А12 или его аналог AI вводили после региональной ишемии (одновременно с началом реперфузии) в дозе 0,35 мкмоль/кг массы животного внутривенно болюсом; в контроле — такой же объем изотонического раствора натрия хлорида (0,5 мл). Выбор доз основан на результатах работы [3]. В конце опыта для идентификации зоны риска и интактной области миокарда реокклюдировали ПНА и в яремную вену вводили болюсно 2 мл 1% раствора Evans Blue. Затем вырезали сердце, выделяли и замораживали ЛЖ для последующего определения размеров ИМ. Замороженный ЛЖ разрезали перпендикулярно длинной оси сердца на 4–5 срезов толщиной около 1,5–2 мм, которые затем инкубировали в течение 10 мин в 1% растворе 2, 3, 5-трифенилтетразолий хлорида в 0,1 М калий-фосфатном буфере (рН 7,4 при 37 °С). Полученные образцы сканировали, площади ИМ и зоны риска определяли методом компьютерной планиметрии, используя программу Imagecal. После этого срезы взвешивали для определения массы ЛЖ. В каждой группе рассчитывали отношение зоны риска/вес ЛЖ (зона риска/ЛЖ) и ИМ/зона риска в процентах [3]. В отдельных сериях опытов ткань зоны риска выделяли в исходном состоянии или в конце реперфузии для определения активности ГП, Сu,Zn-CОД, каталазы и содержания МДА.
Повреждение мембран кардиомиоцитов. В опытах in vivo повреждение мембран оценивали по увеличению активности лактатдегидрогеназы (ЛДГ) и фракции МВ креатинкиназы (MB-КK) в плазме крови. Около 0,5 мл крови собирали в гепаринизированные пробирки из венозного катетера в исходном состоянии (перед окклюзией ПНА) и после часа реперфузии. Активность ферментов в плазме определяли на спектрофотометре Shimadzu UV-1800 при λ=340 нм, используя наборы фирмы «BioSystems».
Активность антиоксидантных ферментов и содержание МДА в сердце. Активность антиоксидантных ферментов определяли в супернатанте 800 g × 10 мин гомогенатов нативного миокарда или зоны риска сердца на регистрирующем спектрофотометре Hitachi-557 при температуре 20 °С. Содержание белка в супернатанте миокарда определяли по методу Лоури.
Активность каталазы определяли по скорости утилизации пероксида водорода при 240 нм [12]. За единицу активности принимали количество фермента, необходимое для восстановления 1 мкмоль Н2О2 в 1 мин на 1 мг белка (коэффициент молярной экстинкции Н2О2=43,6 М-1·cм-1).
Активность Сu, Zn-CОД определяли, согласно методу [13], по ингибированию восстановления синего нитротетразолия супероксидным радикалом, генерируемым в системе ксантин-ксантиноксидаза, определяя кинетику образования формазана при 560 нм. За единицу активности СОД принимали количество фермента, необходимое для 50% подавления восстановления синего нитротетразолия.
Активность Se-содержащей ГП определяли в сопряженной глутатионредуктазной системе по скорости окисления NADPH при 340 нм с гидропероксидом третбутила в качестве субстрата в нашей модификации [14]. Среда определения содержала 0,5 ед/мл активности глутатионредуктазы из дрожжей, 1 мМ GSH, 0,2 мМ гидропероксида третбутила, 0,15 мМ NADPH и 1 мМ ЭДТА в 50 мМ K,Na-фосфатном буфере рН 7,0. При расчете начальной скорости вводили поправку на неферментативное окисление глутатиона за время реакции. За единицу активности ГП принимали количество фермента, необходимое для окисления 1 мкмоль восстановленного глутатиона (GSH) в 1 мин.
Содержание вторичных продуктов свободнорадикального окисления липидов (МДА) в гомогенате миокарда определяли по реакции с 2-тиобарбитуровой кислотой в кислой среде, анализируя количество образовавшегося триметинового комплекса на спектрофотометре Hitachi 220А при 532 нм [15].
Регистрация АФК в перфузате. Оттекающий от перфузируемого сердца крысы перфузат собирали в течение 20-й минуты исходного состояния и 1, 3 и 5-й минут реперфузии в охлажденные льдом пробирки. После добавления к аликвотам перфузатов спиновой ловушки 5,5-диметил-1-пирролин-N-оксид (ДМПО) до конечной концентрации 100 мМ их замораживали и хранили в жидком азоте до регистрации спектров электронного парамагнитного резонанса (ЭПР). Спектры ЭПР регистрировали на спектрометре Х-диапазона типа Е-109Ена уровне СВЧ-мощности 10 мВт; частота СВЧ-поля спектрометра составляла 9,15 ГГц. Сканирование магнитного поля осуществляли в области g=2,0028 [16]. Для количественного определения содержания ДМПО-OH в образцах проводи-ли двойное интегрирование ЭПР-спектров перфузата. Полученные значения сравнивали с аналогичными величинами сигналов водных растворов спиновых зондов ТЕМПО и ТЕМПОЛ.
Статистическая обработка. Приведенные показатели выражены как среднее ± ошибка среднего. Различия между группами оценивали с помощью t-критерия Стьюдента и критерия Ньюмена—Кейлса. Их считали статистически значимыми при р<0,05.
Результаты
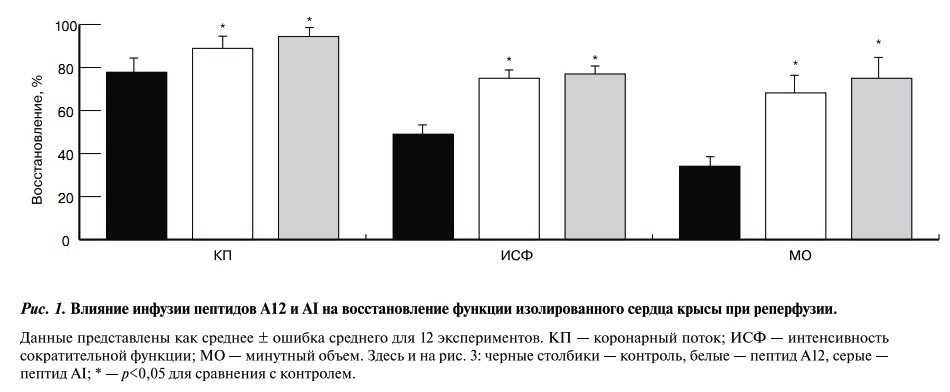
Восстановление коронарного потока и функции сердца при реперфузии.В исходном состоянии (до моделирования глобальной ишемии и реперфузии) коронарный поток перфузируемого сердца составил 18±2 мл/мин, развиваемое ЛЖ давление 99±1 мм рт. ст., частота сокращений сердца 303±2 уд/мин, ИСФ 31380±560 мм рт. ст./мин, минутный объем 44±1 мл. К окончанию реперфузии восстановление коронарного потока, ИСФ и МО в контроле были снижены по сравнению с исходным состоянием (рис. 1).
Рис. 1. Влияние инфузии пептидов А12 и АI на восстановление функции изолированного сердца крысы при реперфузии
Инфузия А12 перед ишемией восстанавливала коронарный поток до 89±5% от исходного и увеличивала восстановление ИСФ и МО соответственно в 1,5 и 2 раза по сравнению с контролем. Сходное влияние на восстановление этих показателей отмечено при введении аналога апелина-12 АI перед ишемией.
Ограничение размеров ИМ. В контроле отношения зона риска/ЛЖ и ИМ/зона риска в конце реперфузии составили 38,6±1,6 и 40,5±2,1% соответственно (табл. 1). Внутривенное введение пептидов А12 или АI не влияло на величину отношения зона риска/ЛЖ, но снижало процентное отношение ИМ/зона риска на 40 и 30% по сравнению со значением в контроле соответственно (p<0,01). Достоверных различий в размерах ИМ между группами А12 или АI обнаружено не было, таким образом, оба пептида в равной степени уменьшали гибель кардиомиоцитов.
Таблица 1. Влияние внутривенного введения пептидов А12 и АI на размер ИМ и активность маркеров некроза в плазме крови у наркотизированных крыс
Активность маркеров некроза в плазме.В исходном состоянии активность MB-КK составляла 274,3±27,2 МЕ/л (см. табл. 1). К концу реперфузии в контроле активность MB-КK возрастала в 7,5 раза по сравнению с исходной. Введение А12 или его аналога АI существенно снижало активность фермента (до 915,1±131,2 и 1200,1±110,1 МЕ/л соответственно; p<0,05) по сравнению с контролем. Активность ЛДГ в плазме в исходном состоянии составляла 91,7±20,5 МЕ/л. В контроле к окончанию реперфузии этот показатель возрастал более чем на порядок. Под действием введения пептида А12 или АI активность ЛДГ в плазме достоверно снижалась в среднем на 47% по сравнению с контролем.
Образование АФК во время ранней реперфузии. Об интенсивности образования АФК в сердце во время ранней реперфузии судили исходя из амплитуды сигнала ДМПО-OH в перфузате. В спектрах ЭПР-перфузатов, полученных во время реперфузии, регистрировали появление 4 узких эквидистантных компонентов с соотношением интенсивностей, равным 1:2:2:1, соответствующих спиновому аддукту ДМПО-OH — соединению, образующемуся при взаимодействии молекул ДМПО и короткоживущих токсичных гидроксильных радикалов (рис. 2). Источниками последних в миокардиальном оттоке могли быть продукты реакций Габера— Вейса и Фентона, а также спонтанный распад нестабильного аддукта ДМПО-ООН, образующегося в результате взаимодействия ДМПО и супероксидных радикалов [16].
Рис. 2. Спектр ЭПР-перфузата изолированного сердца крысы после добавления ДМПО. Показано образование спинового аддукта ДМПО-ОН, сигнал ЭПР которого характеризуется фактором спектроскопического расщепления g=2,0028
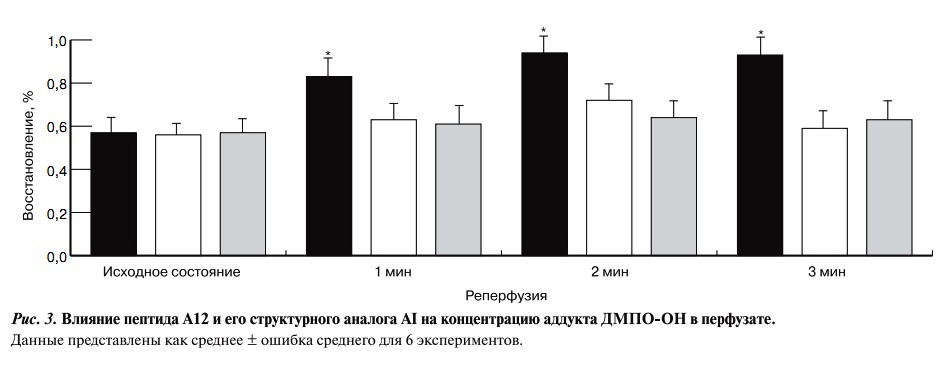
В исходном состоянии во всех группах наблюдалась сопоставимая концентрация аддукта ДМПО-ОН в миокардиальном оттоке (рис. 3).
Рис. 3. Влияние пептида А12 и его структурного аналога АI на концентрацию аддукта ДМПО-ОН в перфузате
В контроле при реперфузии концентрация аддукта ДМПО-ОН существенно возрастала — на 3-й и 5-й минутах реперфузии она была в среднем в 1,6 раза выше, чем до ишемии. Введение перед ишемией A12 или AI достоверно уменьшало образование аддукта ДМПО-ОН по сравнению с контролем. В этом случае концентрация ДМПО-ОН на стадии ранней реперфузии не отличалась от значений в исходном состоянии в этих группах. Эти данные свидетельствуют о снижении выведения из сердца под действием пептидов систем, генерирующих образование АФК, способных вызывать необратимые повреждения кардиомиоцитов. Введение А12 совместно с L-NAME не оказывало достоверного влияния на продукцию ДМПО-ОН во время реперфузии по сравнению с действием одного А12.
Активность СОД, каталазы и ГП, содержание МДА в сердце. В контроле тотальная ишемия и последующая реперфузия изолированного сердца крысы приводили к достоверному увеличению содержания МДА в миокарде к концу реперфузии по сравнению с исходным состоянием (табл. 2). Одновременно отмечена тенденция к снижению активности Сu, Zn-CОД и каталазы в реперфузированном сердце по сравнению с этими показателями в исходном состоянии. Инфузия А12 перед ишемией увеличивала активности Сu, Zn-CОД и каталазы в сердце к окончанию реперфузии в среднем в 1,4 раза по сравнению с контролем, но не влияла на активность ГП. В серии опытов с А12 не отмечено достоверного снижения МДА в миокарде в конце реперфузии по сравнению с контролем. В контроле под действием региональной снижению активности Сu, Zn-CОД и каталазы в реперфузированном сердце по сравнению с этими показателя-ми в исходном состоянии. Инфузия А12 перед ишемией увеличивала активности Сu, Zn-CОД и каталазы в сердце к окончанию реперфузии в среднем в 1,4 раза по сравнению с контролем, но не влияла на активность ГП. В серии опытов с А12 не отмечено достоверного снижения МДА в миокарде в конце реперфузии по сравнению с контролем. В контроле под действием региональной ишемия и возобновления коронарного кровотока к окончанию реперфузии в зоне риска увеличивалось содержание МДА и снижалась активность ГП (p<0,05) и Сu, Zn-CОД (p=0,08) по сравнению с этими показателями в исходном состоянии. Достоверных изменений в активности каталазы в конце реперфузии по сравнению с исходным состоянием не наблюдали. Внутривенное введение А12 после периода региональной ишемии достоверно увеличивало активность ГП в зоне риска в конце реперфузии до значения в исходном состоянии, а активность Сu, Zn-CОД и каталазы — в 1,5 раза по сравнению с контролем. Этот эффект сопровождался снижением содержания МДА в зоне риска до исходного.
Таблица 2. Влияние апелина-12 на активность ферментов антиоксидантной защиты и содержание МДА в изолированном сердце крысы и в зоне риска сердца наркотизированных крыс
Обсуждение
Впервые подтверждена способность С-концевого фрагмента адипокина апелина А12 и его структурного аналога AI уменьшать образование АФК при реперфузии сердца. На это указывает снижение образования аддукта гидроксильных радикалов ДМПО-ОН в перфузате изолированного сердца крысы на первых минутах реперфузии (см. рис. 3).
Учитывая возможность перехвата АФК спиновой ловушкой, а, следовательно, проявление ею кардиозащитных свойств [17], в данной работе мы вводили ДМПО в мио-кардиальный отток. Это ограничение не дает однозначного представления об уменьшении генерации АФК в дыхательной цепи митохондрий под действием ксантиоксидазной системы или в результате перекисного окисления липидов в ткани миокарда. Поэтому в дальнейшем планируется изучение влияния пептидов А12 и АI на продукцию АФК — гидроксил-радикала (ОН) и супероксидного анион-радикала (О2-) — в зоне риска сердца крыс при реперфузии методом ЭПР с помощью микродиализа [18]. Данный подход позволит исключить артефакты, связанные с воздействием самой ловушки на ткань сердца. Тем не менее обнаруженное нами снижение концентрации спинового аддукта ДМПО-OH в перфузате свидетельствует о подавлении пептидами образования высокотоксичных кислородных радикалов, вызывающих необратимые повреждения сердечной мышцы.
Принципиально важным является обнаружение влияния пептида А12 на активность антиоксидантных ферментов — СОД, каталазы и ГП в миокарде при репер-фузии. На обеих моделях ишемического и реперфузионного повреждения сердца (в бóльшей степени у крыс in vivo) введение А12 обеспечивало более эффективную антиоксидантную защиту реперфузированного миокарда, предотвращая снижение или увеличивая активность этих ферментов по сравнению с контролем (см. табл. 2). Повышению компенсаторных возможностей эндогенной антиоксидантной системы в сердце крыс in situ под действием А12 соответствовало снижение содержания вторичного продукта свободнорадикального окисления липидов — МДА. Эти результаты хорошо согласуются с меньшими повреждениями мембран кардиомиоцитов, которые были оценены по активности ЛДГ и МВ-КК в плазме крови крыс при реперфузии (см. табл. 1).
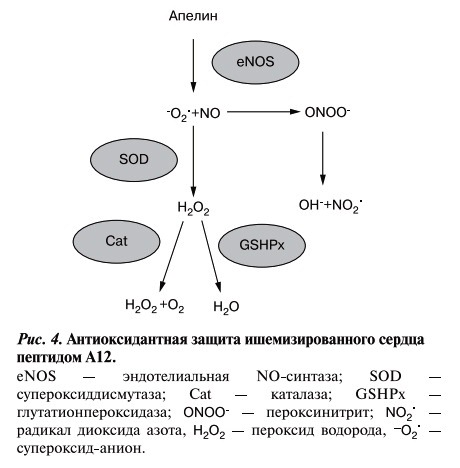
Можно предполагать, что увеличение активности COД под действием A12 приводит к удалению избытка О2-, образующегося в результате ишемического и реперфузионного повреждения сердца (рис. 4). Следствием этого является уменьшение продукции цитотоксичного пероксинитрита (ONOO-) во время ранней реперфузии и, таким образом, окислительного стресса. Образующийся Н2О2, также способен инициировать повреждения миокарда [19], может утилизироваться под действием каталазы и ГП соответственно. Поскольку NO, образующийся под действием А12, и СОД конкурируют между собой за О2-, отсутствие усиления окислительного стресса за счет дополнительного образования ONOO- может быть связано с возросшей активностью СОД.
Рис. 4.Антиоксидантная защита ишемизированного сердца пептидом А12.
Заключение
Снижение продукции короткоживущих активных форм кислорода благодаря улучшению антиоксидантного статуса ишемизированного сердца при реперфузии является одним из принципиальных механизмов кардиозащиты природным пептидом А12 и его модифицированным аналогом АI. Полученные результаты обосновывают целесообразность изучения влияния этих соединений на интенсивность свободнорадикальных процессов в сердце при различной экспериментальной патологии.
Работа выполнена при поддержке гранта РФФИ № 11-04-00078-а.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}