
Защита миокарда от последствий ишемии-реперфузии остается одной из центральных проблем кардиологии. Хорошо известно, что при этом основным фактором повреждения является окислительный стресс. Он возникает в ситуациях, когда антиоксидантная защита кардиомиоцитов не справляется с резко возросшим количеством активных форм кислорода. Во многих экспериментальных работах показано защитное действие различных антиоксидантов при их предварительном введении [1-7]. Однако большинство из них являются синтетическими продуктами, оказывают нежелательное побочное действие, что не позволяет рассматривать их в качестве перспективных терапевтических средств.
Более перспективное направление связано с использованием естественных метаболитов, повышенное содержание которых в кардиомиоцитах способно оказать защитное действие. Примерами могут быть аминокислоты аспартат и глутамат [8], а также их комбинация с динитрозильными комплексами железа [9], обладающими антиоксидантным свойствами [10]. В экспериментах на крысах с кратковременным пережатием коронарной артерии показана способность этой смеси ограничивать размер инфаркта миокарда [9]. Глутамат наряду с другими аминокислотами — цистеином и глицином — входит в состав глутатиона — главного редокс-буфера в кардиомиоцитах. Глутатион находится в клетках в высокой концентрации (5 мМ), что позволяет ему восстанавливать любую дисульфидную связь, образующуюся между цистеинами цитозольных белков. При этом восстановленная форма глутатиона GSH превращается в окисленную GSSG. Обычно соотношение восстановленной формы глутатиона и окисленной ([GSH]/[GSSG]) превышает 10, и поддержание такого соотношения важно для антиоксидантной защиты клеток. Гипоксия или ишемия снижают это соотношение, при этом повышается уровень миоплазматического Са2+ [11]. Транспорт глутатиона в клетки относительно медленный, поэтому лучше использовать моноэтиловый эфир глутатиона — этилглутатион. Он эффективно транспортируется во многие клетки организма, что установлено при введении меченого эфира мышам [12]. Эта способность этилглутатиона была использована при гипоксииреоксигенации изолированного сердца [13]. Сердца крыс подвергали тотальной гипоксии в течение трех 5-минутных периодов, разделенных 5-минутными промежутками реоксигенации. Во время периодов гипоксии в перфузат добавляли этилглутатион (10 мМ), что сопровождалось повышением степени восстановления индекса работоспособности и уменьшением подъема диастолического давления во время гипоксии. При этом общее содержание тиолов в кардиомиоцитах не увеличивалось, и это позволяет предполагать, что защитное действие тиолов во время гипоксии связано с усилением восстановленности тиоловых групп в мембранных белках. Эти работы показали перспективность глутатиона как защитного средства, однако явные их недостатки — кратковременность гипоксии, а также использование очень высокой концентрации этилглутатиона – оставляли открытым вопрос о целесообразности дальнейшей разработки данного соединения в качестве потенциального терапевтического средства.
Целью нашей работы было испытание этилглутатиона в гораздо меньшей концентрации при длительной гипоксии и реоксигенации изолированного сердца.
Методика исследования
Для работы на изолированных сердцах крыс применена стандартная методика ретроградной перфузии [14]. У крыс, анестезированных кетамином (100 мг/кг), вскрывали грудную клетку, извлекали сердце, отмывали его от содержащейся в нем крови. Затем в аорту вводили металлическую канюлю, соответствующую по диаметру аорте, и перфузировали коронарные сосуды стандартным раствором Кребса—Хензелайта с добавлением 11 мМ глюкозы. Раствор предварительно насыщали карбогеном (95% О2 и 5% СО2). В левый желудочек сердца вводили латексный баллончик, заполненный таким количеством жидкости, чтобы диастолическое давление в нем составляло 10—15 мм рт. ст., при котором достигается оптимальное растяжение камеры желудочка. Эта ретроградная перфузия проходила под постоянным давлением 70 мм рт. ст. в течение 20 мин; в это время происходила стабилизация работы сердца. На правое предсердие накладывали электроды для электростимуляции. Перфузия происходила в специализированной установке Hugo Saks.
Для регистрации давления в баллончике, электрограммы и скорости коронарной перфузии использовали датчики и усилители Harward Instruments, аналого-цифро-вой преобразователь (АЦП: USB-6215, используемая частота оцифровки 1000 Гц) и персональный компьютер. Регистрация сигналов происходила непрерывно на протяжении всего опыта.
Управление работой АЦП и передачей сигналов в компьютер, а также запись первичных сигналов в виде бинарных файлов на жесткий диск компьютера, вывод сигналов на экран монитора, их предварительная обработка непосредственно во время эксперимента, введение меток воздействий во время эксперимента, предустановка интервалов изменения режимов стимуляции (при навязывании частоты возбуждений) и возможность последующего анализа как расчетных параметров зарегистрированных сигналов, так и самих этих сигналов, обеспечивались специально созданными нами компьютерными программами (использована графическая среда программирования LabVIEW).
Обработка сигналов непосредственно во время проведения исследования предусматривала расчет функционально значимых параметров в каждом кардиоцикле (по сигналу давления в желудочке определяются частота его сокращений, максимальное, конечное диастолическое и развиваемое давление, а также максимальные скорости нарастания и спада давления; для остальных сигналов рассчитывали их среднюю величину), усреднение этих параметров за каждые 2 с, запись всех расчетных параметров (как по каждому кардиоциклу, так и усредненных за 2 с) в текстовые файлы и непрерывный вывод усредненных значений в виде трендов на экран монитора.
На основе измеренных величин рассчитывали индекс работы сердца (произведение развиваемого давления и частоты сокращений), характеризующий общий расход энергии, а также индекс расслабления (частное от деления максимальной скорости падения давления на величину развиваемого давления). Анализировали качественный состав аритмий; помимо общего количества экстрасистол, учитывали также групповые экстрасистолы. Для оценки степени тяжести аритмии вычисляли интегративный индекс аритмии. Для его расчета тяжесть аритмии выражали в баллах: одиночные экстрасистолы — 1 балл, бигеминия — 2 балла, сальво (строенные экстрасистолы) — 3 балла. Частота идиовентрикулярного ритма колебалась от 6,5 до 11 Гц, в качестве средней величины была избрана 8 Гц (8 баллов). Период фибрилляции расценивали как 20 баллов. Умножая эпизоды аритмии (баллы) на их продолжительность (с) и суммируя эти величины для каждого опыта, получали величину индекса аритмии, а разделив ее на длительность периода наблюдения, получали среднюю величину индекса за минуту.
Протокол опыта: после стабилизации показателей функции осуществляли ступенчатую электростимуляцию с частотой, превышающей спонтанную, с шагом 0,5 Гц вплоть до 11,5 Гц по 15 с на каждой частоте. Через 10 мин начинали гипоксическую перфузию (30 мин) посредством смены перфузата, насыщенного карбогеном, на перфузат, не содержавший кислорода (95% азота и 5% СО2). Затем возобновляли аэробную перфузию также на 30 мин.
На заключительном этапе опыта вновь выполняли пробу с навязыванием повышенной частоты возбуждений.
Результаты
Исходные величины функциональных показателей сердца имеют большое значение для степени восстановления функции после гипоксии-реоксигенации. Известно, что чем ниже интенсивность окислительного метаболизма к началу гипоксии, тем лучше степень восстановления после реоксигенации. В связи с этим в обработку брали только опыты с развиваемым давлением не ниже 130 мм рт. ст. и частотой сокращений не ниже 230 в минуту.
Гипоксическая перфузия сердца сопровождалась крутым снижением развиваемого давления уже в первые минуты гипоксии, при этом в опытах без глюкозы она снижалась круче. В этих опытах сократительная активность желудочков прекращалась уже через 10 мин, в то время как при наличии глюкозы она сохранялась на уровне 22—25 мм рт. ст. в периоде 10—30 мин. Отмечена также заметная разница по реакции диастолического давления, характеризовавшего контрактуру миокарда.
Наличие глюкозы в перфузате заметно снижало величину гипоксической контрактуры (рис. 1).
Рис. 1. Динамика диастолического давления (ДАД) в желудочке (М±m) во время гипоксии (0-30 мин) и реоксигенации (31-60 мин) в контрольных опытах с наличием (темные квадраты, Г=11) и в отсутствие (светлые квадраты, Г=0) глюкозы в гипоксическом перфузате. Стрелка указывает начала реоксигенации.
Крутизна подъема диастолического давления начинала достоверно различаться уже со 2-й минуты, и это различие (около 20 мм рт. ст.) сохранялось на протяжении всего гипоксического периода. В периоде реоксигенации контрактура в обеих сериях уменьшалась, причем в серии без глюкозы это происходило более круто, и разница между сериями нивелировалась. Добавление этилглутатиона (0,2 мМ) в гипоксический раствор, содержавший глюкозу, никак не повлияло на динамику гипоксической и реоксигенационной контрактуры (данные не приведены для сохранения ясности рисунка).
Различие между сериями было заметно и при сравнении динамики частоты сокращений. В серии без глюкозы частота сокращений круто падала в 2 раза уже через 3 мин, в то время как при наличии глюкозы — через 5-6 мин. В гипоксическом периоде при наличии глюкозы наблюдали аритмии, которые резко усиливались при реоксигенации. Экстрасистолии, преимущественно желудочкового происхождения, — нередкое явление при работе изолированного сердца в изоволюмическом режиме. Они обусловлены механическим раздражением эндокарда левого желудочка раздутым латексным баллончиком. Детальный анализ аритмии во время реоксигенации (табл. 1) показал, что среднее число таких экстрасистол за 1 мин в группе этилглутатиона в период реоксигенации приблизительно соответствовало величине в исходном состоянии, в то время как в контрольной группе оно было достоверно выше почти в 4 раза. Желудочковую тахикардию или идиовентрикулярный ритм наблюдали во всех 11 опытах контрольной группы, а фибрилляцию — в 9 опытах. В группе этилглутатиона эпизоды желудочковой тахикардии и фибрилляции были в 3 опытах из 9.
Число эпизодов идиовентрикулярного ритма и фибрилляции в группе этилглутатиона было соответственно в среднем в 19 и 4 раза меньше по сравнению с контрольной группой (см. табл. 1). Известно, что фибрилляция сердец крыс может спонтанно прекращаться, поэтому в табл. 1 включены также данные по продолжительности этих периодов. Средняя длительность периодов идиовентрикулярного ритма и фибрилляции в группе этилглутатиона была в 7-12 раз меньше, чем в контрольной группе (см. табл. 1).
Таблица 1. Характеристика аритмии во время реоксигенации при действии этилглутатиона (гипоксия с глюкозой)
Наконец, величина интегративного индекса аритмии, включающая все виды аритмий, а также их длительность, была в 5 раз (p<0,01) меньше в группе этилглутатиона.
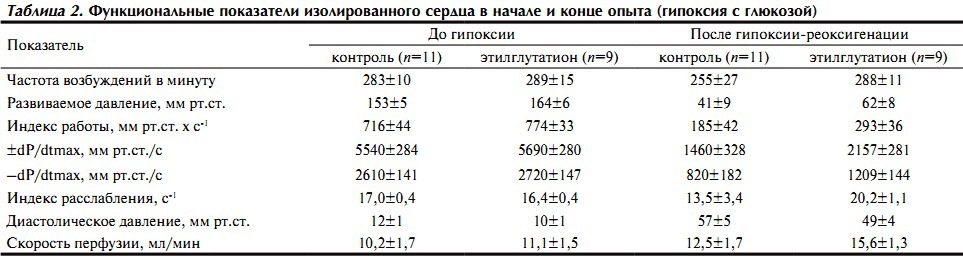
К концу реоксигенационного периода в контрольной группе в 3 опытах из 11 не наблюдали восстановления сократительной активности желудочка, а при добавлении этилглутатиона она восстанавливалась во всех опытах. Это сопровождалось лучшим восстановлением развиваемого давления, а также индекса работы сердца — его величина превышала контрольный уровень на 58% (p=0,06). Кроме того, примерно в 1,5 раза была выше максимальная скорость сокращения и расслабления (табл. 2). В обеих группах скорость коронарной перфузии умеренно возрастала по сравнению с исходным уровнем, что в условиях постоянного перфузионного давления свидетельствует о снижении тонуса коронарных сосудов, особенно если учесть, что часть сосудов не перфузировалась из-за контрактуры миокарда.
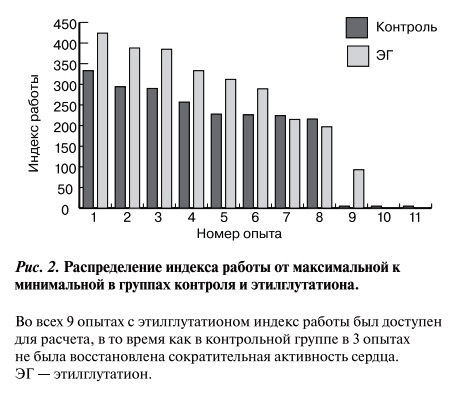
Высокие средние ошибки показателей после реоксигенации обусловлены большой вариативностью опытов. В этой ситуации вполне оправдано сравнение групп по отдельным опытам. Распределение величин в каждой группе от максимальной к минимальной (рис. 2) свидетельствует, что 7 величин в группе этилглутатиона выше, чем в контрольной группе, и лишь в 2 опытах величины примерно равны. Для уравнивания числа опытов в группах из контрольной группы были исключены две вели-чины — максимальная и минимальная. Это позволило оценить разницу по парному критерию Стьюдента: он показал очень высокую степень достоверности (p<0,001). Этот результат позволяет полагать, что степень восстановления индекса работы в группе этилглутатиона достоверно выше, чем в контрольной группе.
Рис. 2. Распределение индекса работы от максимальной к минимальной в группах контроля и этилглутатиона
В отсутствие глюкозы в гипоксическом перфузате аритмии были редкими, а степень восстановления развиваемого давления к концу реоксигенации практически не отличалась от представленных в табл. 2 контрольных данных или немного превышала их. Так, в серии с глюкозой степень восстановления развиваемого давления составляла 26±5%, а в опытах без глюкозы — 36±3% (различия недостоверны). Добавление этилглутатиона к гипоксическому перфузату без глюкозы (n=5) значительно ухудшило восстановление функции — степень восстановления развиваемого давления составила 18±4%, т.е. в 2 раза ниже контрольной величины. Таким образом, этилглутатион улучшал восстановление функции только при наличии глюкозы в гипоксическом перфузате.
Таблица 2. Функциональные показатели изолированного сердца в начале и конце опыта (гипоксия с глюкозой)
Через 10 мин после завершения 30-минутного реоксигенационного периода все сердца подвергали электростимуляции с постепенно возрастающей частотой. К этому времени в контрольной группе появилась сократительная активность желудочков в 2 опытах из 3, у которых она отсутствовала к концу реоксигенации, так что электростимуляция была выполнена в 10 контрольных опытах. Однако в 2 опытах навязать повышенную частоту не удалось, несмотря на 4-кратное усиление вольтажа сверх порогового уровня. Кроме того, в 4 опытах максимально воспроизводимой частотой стала 480—510 в минуту, и только в двух опытах была воспроизведена максимальная частота стимуляции — 690 в минуту. Добавление этилглутатиона оказало положительное действие — лишь в 4 опытах максимально воспроизводимой частотой стала 450—510 в минуту, а в 7 опытах была достигнута максимальная частота. Таким образом, максимальная частота стимуляции была воспроизведена в 22% контрольных опытов и в 64% опытов с добавлением этилглутатиона. В среднем максимально воспроизводимой частотой в контрольной серии стала 513±48 в минуту, а в серии с этилглутатионом максимально воспроизводимая частота составляла 611±33 в минуту, т.е. на 23% выше. Различия достоверны при сравнении по парному критерию Вилкоксона.
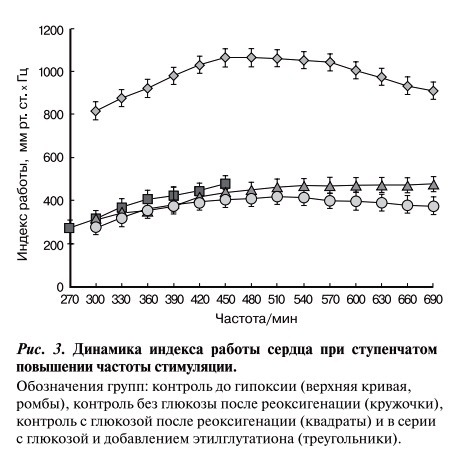
Повышение частоты возбуждений, как правило, сопровождалось постепенным уменьшением развиваемого давления. Однако степень этого снижения на начальном этапе стимуляции была меньше, чем степень повышения частоты, поэтому индекс работы сердца (произведение давления и частоты) повышался и лишь затем немного снижался (рис. 3, верхняя кривая). После гипоксии-реоксигенации в контрольных опытах данная зависимость рассчитана лишь для опытов, воспроизводивших частоту не более 450 в минуту. Как видно из рисунка, индекс работы сердец, получавших этилглутатион, даже при максимальной частоте стимуляции был выше, чем при начальной частоте, подобно тому, как это наблюдалось до гипоксии. В контрольных опытах без глюкозы в гипоксическом перфузате способность к воспроизведению высокой частоты полностью сохранялась (см. рис. 3).
Рис. 3. Динамика индекса работы сердца при ступенчатом повышении частоты стимуляции
Обсуждение
Результаты работы могут быть суммированы в виде двух основных положений: 1) наличие глюкозы в гипоксическом перфузате сопровождается тяжелыми аритмиями при реоксигенации и нарушением воспроизведения высокой частоты стимуляции; 2) этилглутатион в этих условиях предотвращал или значительно уменьшал эти функциональные нарушения, причем использованная дозировка препарата была в 50 раз меньше, чем в опытах на том же объекте [13].
Наличие глюкозы в перфузате, что соответствует естественной ситуации, способствует поддержанию сократительной и электрической активности сердца, хотя и на низком уровне, а также уменьшает падение уровня АТФ в клетках во время гипоксии [15]. Эта ситуация характеризует противоречие между реакцией сердца как органа и функцией сердца как части организма. При гипоксии органа лучшим способом его выживания является состояние гибернации, сочетающееся с распадом гликогена, что характерно для ишемии. Для организма необходимо поддержание функции сердца, хотя бы и в ослабленном виде, и наличие глюкозы как раз служит этой цели, нарушая естественную защитную реакцию кардиомиоцитов и органа в целом. Полученные данные позволяют предполагать, что наличие глюкозы в перфузате при гипоксии или при периодической перфузии ишемического сердца не всегда является необходимым фактором сохранения жизнеспособности миокарда.
Появление аритмий в этот период, скорее всего, обусловлено гетерогенностью состояния кардиомиоцитов в различных участках желудочков, вызванной сильной контрактурой миокарда и обусловленным ею пережатием сосудов в некоторых участках. Хотя при наличии глюкозы в перфузате величина контрактуры меньше, чем в отсутствие глюкозы, гетерогенность состояния кардиомиоцитов, несомненно, больше, потому что постоянный приток глюкозы в перфузируемые участки поддерживает часть кардиомиоцитов в рабочем состоянии и тем самым создает условия для гетерохронного возбуждения разных участков миокарда. Аритмия усиливается еще больше в начале реоксигенации, когда приток кислорода возвращает сократительную способность для большего количества клеток.
Роль гликолиза в обеспечении кальциевого транспорта в кардиомиоцитах предсердий детально рассмотрена в работе [16]. В интактных регулярно сокращающихся миоцитах ингибиторы гликолиза — 2-деоксиглюкоза и йодацетат, не влиявшие на состояние митохондрий, снижали уровень Са2+ в систоле и повышали в диастоле, а также замедляли скорость удаления Са2+ в саркоплазматическую сеть. Эти данные означают, что глюкоза способствует ускорению транспорта Са2+ в саркоплазматическую сеть и выходу из нее; в условиях недостаточного энергоснабжения это провоцирует возникновение аритмий. Аритмогенный эффект глюкозы был показан также на эмбриональных кардиомиоцитах [17], и различные ингибиторы гликолиза — йодацетат, деоксиглюкоза — оказывали заметное защитное действие [18].
Нарушение возбудимости и проводимости желудочков оказалось в некоторых опытах настолько резким, что сердца не смогли восстановить свою сократительную активность даже к концу реоксигенационного периода. Кроме того, более 50% сердец в данной группе не смогли воспроизводить высокую частоту сокращений. Эти изменения свидетельствуют о нарушении системы ионного транспорта, в частности, белков кальциевого транспорта в кардиомиоцитах. Вход Са2+ через сарколемму осуществляется в основном через так называемые медленные кальциевые каналы, а выход — через Na+/ Са2+ обменник. Внутриклеточный уровень Са2+ определяется активностью белков саркоплазматической сети. При этом поглощение Са2+ из миоплазмы происходит благодаря функции Са2+-АТФазы (SERCA2), а выход — через воротный белок, чувствительный к рианодину и названный рианодиновым рецептором (RyR2). Пропускная способность этих белков зависит от редокс-потенциала клеток.
Тиоловые группы цистеина, входящего в состав этих белков, могут подвергаться редокс-модификациям — либо окислению с образованием дисульфидных связей, либо восстановлению путем глутатионилирования или нитрозилирования (ковалентное присоединение NO с образованием SNO). Окисление небольшого числа этих групп повышает чувствительность белков, пропускающих Са2+ в миоплазму, к концентрации Са2+ [19, 20], способствуя учащению и усилению сокращений, одновременно замедляя удаление Са2+ из миоплазмы в саркоплазматическую сеть, но при окислении большого количества этих групп происходит обратный процесс [21]. Задержка Са2+ в саркоплазматической сети уменьшает потребление АТФ и тем самым способствует сохранению энергетического потенциала кардиомиоцитов.
Повышение уровня глутатиона в клетках во время гипоксии, вероятно, ограничивает степень окисления белков кальциевого транспорта и способствует закрытию канала RyR2 [20]. Тем самым уменьшается гетерогенность миокарда, что проявляется, в частности, в лучшей стабилизации ритма сокращений. Однако для такого заключения нужно изучение степени глутатионилирования белков кальциевого транспорта. Косвенным аргументом в пользу этого является неэффективность этилглутатиона в отсутствие глюкозы в гипоксическом перфузате, когда деятельность сердца прекращалась. Следует также отметить, что этилглутатион никак не повлиял на степень гипоксической или реоксигенационной контрактуры, зависимой главным образом от наличных запасов АТФ в кардиомиоцитах. Это соответствует известным данным о способности глутатиона эффективно предотвращать нарушения сократительной функции, вызванные окислительным стрессом, но не гипоксией или ишемией [22-24], при которых больше всего страдает система энергообеспечения.
Таким образом, наблюдавшиеся нами эффекты уменьшения аритмии и лучшего восстановления сократительной функции после реоксигенации под влиянием этилглутатиона могут быть объяснены уменьшением окисления белков кальциевого транспорта. Эффективность этилглутатиона в этих условиях привлекает внимание к редокс-регуляции функции белков кальциевого транспорта как способу предотвращения постишемических нарушений функции сердца.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}