
Микрорибонуклеиновые кислоты (микроРНК) представляют собой класс олигонуклеотидов, в состав которого входят 20—25 нуклеотидов. Их основная функция — торможение трансляции сигнала от информационной РНК (иРНК) в процессе синтеза белков на рибосомах, а также в активации системы деградации иРНК [1, 2]. Эти особенности позволяют микроРНК составлять важные компоненты системы контроля баланса между скоростью пролиферации и дифференцировки клеток в процессе онтогенетического развития органа, а также в процессе канцерогенеза [3, 4]. В настоящее время определены около 600 микроРНК, участвующих в регуляции более 50% генов человека, ответственных за синтез различных факторов, присущих клеточному гомеостазу. Поскольку микроРНК вставлены в интроны генов кодирования или в эксоны, их транскрипция зависит от доминантного гена [5].
Имеются доказательства, что экспрессия микроРНК является сложным процессом, в котором участвуют различные рибонуклеазы и эндонуклеазы, нейроэндокринные факторы, цитокины, кемокины. Результаты многих исследований доказывают участие микроРНК-145 и микроРНК-143 в поддержании равновесия между секреторным (пролиферативным) и сократительным (паттерном клеточной дифференциации) фенотипами гладких мышечных клеток сосудов (ГМКС) — в процессах, которые непосредственно влияют на реактивность сосудов, податливость и эластичность сосудистой стенки [6, 7]. Y. Cheng и соавт. (2009) представили доказательства роли микроРНК-145 в формировании неоинтимы в сосуде после его повреждения, и это согласуется с гипотезой, допускающей микроРНК-145 в качестве маркера фенотипа ГМКС [8].
Примечательны в этом отношении исследования L. Elia и соавт. (2009), проведенные на трансгенных мышах knockout, по изучению микроРНК-143 и микроРНК-145. В ходе этих исследований определено нарушение сосудистого гомеостаза из-за снижения количества сократительного фенотипа сосудистых миоцитов в результате активации матричных протеиназ и сосудистого ремоделирования, свойственного дисфункции эндотелия [9].
Миокардин — основной ген, управляющий дифференциацией сосудистых миоцитов и индуцирующий экспрессию микроРНК-143 и микроРНК-145, которые могут служить генетическими маркерами ГМКС относительно ее ответа на воздействие инотропных факторов (например, эндотелина-1), образования сократительных миофиламентов и функционирования систем регулирования метаболизма кальция [7].
Важно отметить, что повреждения сосудов ассоциируются со снижением экспрессии микроРНК-145 и ингибицией миокардина вследствие повышенной экспрессии сигнальной молекулы KLF-5 вследствие подавления экспрессии микроРНК-145 [8]. Более того, ключевой фактор, стимулирующий пролиферацию ГМКС, вырабатываемый тромбоцитами (тромбоцитарный β-фактор роста — ТФРβ), был обнаружен в мышиной модели сосудистых повреждений и проявил себя как выраженный ингибитор микроРНК-145. Его экспрессия была подтверждена на сосудистых миоцитах, но не была инди- визуализирована на поверхности эндотелиальных клеток сосудов. Доказана его корреляция с генетическими маркерами сократительных белков (актином, миозином, калпомином), что указывает на значимость гладких миоцитов в биологии, регенерации и реактивности сосуда.
Следует отметить, что как ТФРβ, так и ГМКС участвуют в патогенезе формирования и пролиферации неоинтимы, которая является важным звеном в развитии рестеноза коронарной артерии (КА) после ангиопластики, в том числе с использованием внутрикоронарного стента [10]. Коронарная мышечная клетка становится мишенью для пролиферативного и провоспалительного действия цитокинов, освобожденных под стимулирующим влиянием ТФРβ. Миграция и пролиферация сосудистых миоцитов, способствующие формированию неоинтимы, — процессы, свойственные секреторному фенотипу миоцита, который подвергается митогенному воздействию различных циркулирующих факторов (цитокины, эндотелин-1, ангиотензин II) [11]. Таким образом, система контроля и регулирования соотношения секреторного и сократительного фенотипов ГМКС играет определяющую роль в гиперплазии неоинтимы, которая поддерживается избыточной продукцией внеклеточного матрикса и, следовательно, непосредственно влияет на риск развития рестеноза после ангиопластики.
Исходя из изложенного была поставлена цель работы: определить экспрессию микроРНК-143 и микроРНК-145 в тканевом содержимом внутристентового рестеноза, а также ее соотношение с содержанием коллагена типа I и коронарных ГМКС с контрактильным фенотипом.
Материал и методы
В образцах ткани, взятых из рестенозированных стентов у 5 умерших пациентов, была определена экспрессия микроРНК-143 и микроРНК-145 с использованием морфологических методов [6]. Степень внутристентового рестеноза гистологически была классифицирована как низкая, средняя и выраженная. Образцы тканей изучали методом гибридизации in situдля выявления микроРНК-143 и микроРНК-145 [12, 13], а антитела к α-актину ГМКС, к фибриллярному коллагену типа I или к денатурированному коллагену I применяли для идентификации миоцитов с контрактильным фенотипом и деградации коллагена. Образцы тканей были изучены с помощью конфокальной и иммунофлюоресцентной микроскопии, методов Western blot и Northen blot.
Морфологические исследования были проведены в Лаборатории молекулярной и структурной биологии Института сердца и легких Max-Planck (Bad Nauheim, Германия).
Результаты
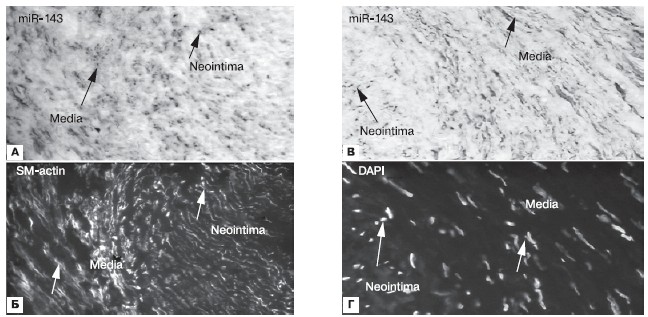
Прежде всего следует отметить релевантность метода гибридизации in situдля определения микроРНК в ткани, взятой из рестенозированного стента. В зонах медии и интимы рестенозированной КА обнаружена экспрессия микроРНК-143 (рис. 1, А). Кроме того, в медии выявлен актин — один из сократительных белков гладких миоцитов (рис. 1, Б).
Рисунок 1.Релевантность метода гибридизации in situдля определения микроРНК в ткани, взятой из рестенозированного стента.
А — экспрессия микроРНК-143 в медии и неоинтиме рестенозированной артерии; Б — содержание актина в медии рестенозированной артерии; В — гаадкий миоцит в интиме коронарной артерии при рестенозе; Г — подтверждение методом DAPI наличия клеточных ядер.
Чтобы подтвердить достоверность данных, полученных с помощью метода индивидуализации микроРНК в ткани рестеноза, и концептуальное значение этого доказательства, указываем на то, что в ходе использованного in situтеста гибридизации отсутствуют артефакты. Иммунофлюоресцентный метод позволяет четко определять клеточные ядра, меченные DAPI, которые свидетельствуют о целостности клеток тканевого паттерна рестеноза (рис. 1, Г). Используя праймеры к микроРНК-143, мы идентифицировали гладкие миоциты в интиме рестенозированного стента (рис. 1, В). Важно отметить, что мышечные клетки неоинтимы меньше в размерах, чем миоциты из зоны медии КА.
Полученные данные показывают, что по мере развития внутристентового рестеноза экспрессия микроРНК-143 уменьшается.
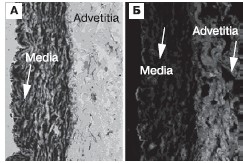
В отсутствие рестеноза выявили значительную экспрессию микроРНК-143, что сочеталось с содержанием в медии КА большого числа гладких миоцитов (рис. 2, А). Более того, микроРНК-143 не обнаружено в адвентиции артериальной стенки, что указывает на принадлежность этой семьи микроРНК к биологии ГМКС. Следует также отметить наличие очень незначительного количества (которым можно пренебречь) деградированного коллагена в медии и главным образом в адвентиции КА (рис. 2, Б).
Рисунок 2. Высокая экспрессия микроРНК-143 и большое число миоцитов в медии коронарной артерии (А, стрелка) и незначительное содержание денатурированного коллагена типа I в медии и адвентиции (Б, стрелки) в отсутствие рестеноза.
Примечательно, что уже при умеренной степени рестеноза определяется явное снижение экспрессии микроРНК-143 в зоне медии коронарного сосуда (рис. 3, А). На этом фоне идентифицируется увеличение количества деградированного коллагена (рис. 3, Б). По мере прогрессирования внутристентового рестеноза эти явления становится более выраженными.
Рисунок 3. Снижение в медии экспрессии микроРНК-143 (А, стрелка) и увеличение количества деградированного коллагенатипа I (Б, стрелка) при умеренной степени внутристентового рестиноза.
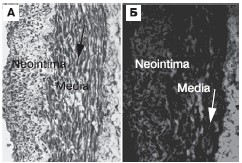
При резком рестенозе в медии и неоинтиме сегмента КА отмечается дальнейшее увеличение числа гладких миоцитов с секреторным фенотипом, которые не экспрессируют микроРНК-143 (рис. 4, А), а также количества деградированного коллагена типа I (рис. 4, Б). Выраженное снижение содержания микроРНК-143 обнаруживается как в медии, так и в неонтиме.
Рисунок 4. Выраженный внутристентовый рестеноз.
Одним из морфологических признаков развития рестеноза является уменьшение соотношения коллагена типа I (фибриллярный) к денатурированному коллагену на фоне количественного снижения микроРНК-143 и коронарных миоцитов с сократительным фенотипом. Степень снижения содержания фибриллярного коллагена типа I и увеличения деградированного коллагена растет в ходе развития рестеноза (рис. 5, см. цв. вклейку). Так, при выраженном внутристентовом рестенозе отмечается массивная деградация коллагена типа I в медии вплоть до его исчезновения, что сопровождается значительным накоплением деградированного коллагена (см. рис. 5, K—М, см. цв. вклейку). Если в нормальном сосуде соотношение между денатурированным коллагеном I и коллагеном I фибриллярным составляет около 0,5, то при выраженной степени рестеноза этот показатель увеличивается в 20 раз, составляя в среднем 12,4.
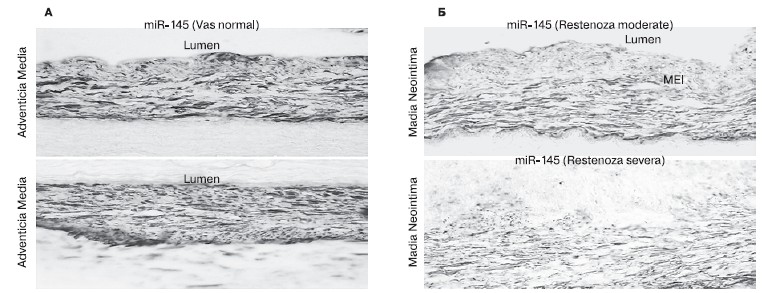
Концептуально важно подчеркнуть, что количественные изменения, подобные микроРНК-143, были обнаружены и по отношению к микроРНК-145 (рис. 6). В медии нормальной КА (см. рис. 6, А) выявляется снижение количества микроРНК-145 уже при умеренной степени рестеноза, которое становится более значительным при резком рестенозе (см. рис. 6, Б).
Рисунок 6. Экспрессия микроРНК-145 в медии нормальной коронарной артерии (А) и снижение экспрессии микроРНК-145 при средней степени и выраженном рестенозе (Б).
Таким образом, развитие внутристентового рестеноза после ангиопластики сопровождается снижением содержания микроРНК-143 и микроРНК-145 в медии КА, что сопровождается явлениями, характерными для формирования неоинтимы: уменьшением числа миоцитов с контрактильным фенотипом и активацией деградации фибриллярного коллагена I типа.
Обсуждение
Один из ключевых механизмов формирования и пролиферации неоинтимы, что представляет собой морфологический субстрат внутристентового рестеноза, связан с дисфункцией эндотелия и нарушением системы контроля фенотипа гладких миоцитов КА [11, 14]. В отличие от нормальных сосудов, в интиме рестеноза появляются коронарные миоциты, уменьшенные в размере и имеющие синтетический фенотип. Миграция мышечных клеток из медии к эндотелию находится под контролем различных локальных систем. Одна из них представлена пассивным барьером внеклеточной матрицы, состоящей из коллагена. Вторая основана на функционировании микроРНК-143 и микроРНК-145, которые поддерживают сократительный фенотип зрелых ГМКС и противостоят действию факторов, способствующих их превращению в секреторных миоцитах, предрасположенных к миграции пролиферации. В этом контексте предполагалось участие микроРНК-143 и микроРНК-145 в патогенезе формирования неоинтимы и внутристентового рестеноза, в связи с чем исследовали их экспрессию в тканевом паттерне рестеноза, взятом у умерших пациентов. Кроме того, мы определяли маркер сократительных миоцитов актина, как и степень деградации коллагена типа I в медии и в неоинтиме. Полученные данные четко указывают на снижение количества микроРНК-143 и микроРНК-145 в прямой зависимости от степени рестеноза. Более того, уменьшению экспрессии микроРНК соответствует снижение содержания актина, а также фибриллярного коллагена I типа; при этом отмечается увеличение содержания деградированного коллагена типа I в медии и интиме. Следовательно, уменьшение экспрессии микроРНК-143 и микроРНК-145 в медии КА может быть рассмотрено как важный механизм приобретения миоцитами синтетического фенотипа и их миграции к интиме — процесс, поддерживаемый деградацией внеклеточного коллагена. Последнее явление может быть, в частности, следствием освобождения мышечными клетками протеиназ (прежде всего коллагеназы, MMP-9), которые приводят к расщеплению фибриллярного коллагена внеклеточного матрикса из различных зон стенки сосуда. Механизмы, которые могут вызвать снижение экспрессии микроРНК-143 и микроРНК-145, остаются невыясненными. Поскольку миграция гладких миоцитов и пролиферация неоинтимы ассоциируются с активацией как местной, так и генерализованной системы ангиотензина, можно предположить роль ангиотензина II в этом процессе. Знание механизмов, которые контролируют экспрессию микроРНК-143 и микроРНК-145, является важным в силу возможного их моделирования различными фармакологическими средствами, что позволит значительно снизить риск развития рестеноза КА после ангиопластики. Перспектива этого подхода определена и тем, что в эксперименте увеличением экспрессии микроРНК-145 под воздействием аденовируса удалось предупредить формирование неоинтимы сонной артерии крысы после баллонной ангиопластики [7]. Другие семьи микроРНК, которые, согласно данным литературы, могут быть вовлечены в процесс рестеноза КА, являются микроРНК-21 и микроРНК-221 [15, 16]. В отличие от регрессивной динамики микроРНК-143 и микроРНК-145, их экспрессия при рестенозе повышена и ассоциируется со степенью пролиферации неоинтимы. Проведены исследования на различных видах животных с целью определения возможности улучшения ремоделирования миокарда и других тканей путем введения веществ миметического действия на микроРНА [16—18]. А. Care и соавт. (2007) опубликовали результаты, указывающие на роль микроРНК-133 в регуляции гипертрофии кардиомиоцитов [19].
Таким образом, нами впервые на клиническом материале установлено уменьшение экспрессии микроРНК-143 и микроРНК-145, которое сопровождается изменением фенотипа гладких миоцитов — снижением экспрессии миоцитов с контрактильным фенотипом и увеличением миоцитов секреторного фенотипа, — явление, которое может быть рассмотрено как важный механизм формирования гиперплазии неоинтимы и развития внутристентового рестеноза КА после ангиопластики. Исследование факторов, определяющих снижение микроРНК-143 и микроРНК-145, представляется важным не только в плане их возможного модулирования с целью снижения риска рестеноза, но и выявления новых предикторов его диагностики и прогнозирования.
Автор выражает искреннюю благодарность проф. Костину С., зав. лабораторией молекулярной и структурной биологии Института сердца и легких Max-Planck (Bad Nauheim, Германия) за содействие в реализации морфологических исследований.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}