
Распространенность ожирения растет как в экономически развитых, так и развивающихся странах, и представляет серьезную медико-социальную проблему, поскольку, с одной стороны, ожирение признано одним из модифицируемых факторов риска развития атеросклероза, а с другой, висцеральная форма ожирения является одним из пусковых механизмов появления инсулинорезистентности и часто сочетается с артериальной гипертензией (АГ), липидными и углеводными нарушениями, повышая риск развития сахарного диабета (СД) 2-го типа [1].
Причинами развития ожирения признаны изменения в образе жизни с преобладанием в рационе рафинированных продуктов, жиров животного происхождения и легкоусвояемых углеводов, а также со снижением физической активности. Однако факторы окружающей среды оказывают более выраженное воздействие у лиц с генетической предрасположенностью. По данным разных исследований, генетические факторы обусловливают от 40 до 70% изменений массы тела [2], в связи с чем ведется активный поиск новых генов, отвечающих за жировой и энергетический обмен в организме.
Одним из регуляторов жирового обмена в организме человека является G-протеин, представляющий собой гетерогенный белок, локализующийся на клеточной мембране и состоящий из α-, β- и γ-субъединиц, от взаимодействия которых зависит активность рецептора. При активации рецептора белок G образует ди-, тримеры, которые могут активировать/ингибировать внутриклеточные процессы: активность ионных каналов, фосфолипазу, аденилатциклазу и другие ферменты [3-5]. Известно несколько генетических полиморфизмов белка G, из которых клинически наиболее значимым является полиморфизм С825Т гена субъединицы β3. Аллель 825Т ассоциируется с экспрессией укороченного, функционально активного димера белка G и усилением внутриклеточных сигналов трансдукции [6]. В клинических исследованиях показана ассоциация этого полиморфизма с предрасположенностью к развитию ожирения и АГ [7-12]. Однако данные разных исследований противоречивы. Изучение генетических маркеров ожирения в киргизской этнической группе ранее не проводилось.
Целью настоящего исследования являлось изучение взаимосвязи полиморфизма С825Т гена субъединицы β3 белка G c наличием ожирения у этнических киргизов.
Материал и методы
В исследование были включены 210 человек, из которых 89 пациентов (54 мужчины, 35 женщин) с ожирением (индекс массы тела — ИМТ ≥30 кг/м2) и 121 сопоставимых по полу и возрасту практически здоровых лиц (83 мужчины, 38 женщин) с нормальной массой тела. В исследование не включали лиц с тяжелыми сердечно-сосудистыми (сердечная недостаточность, мозговой инсульт, острый инфаркт миокарда, тромбоэмболия легочной артерии, тяжелые пороки сердца и т.д.) и соматическими заболеваниями (хронические гепатиты, печеночная, почечная недостаточность и хроническая обструктивная болезнь легких III ст., ревматологические заболевания, системная красная волчанка, ревматоидный артрит и другие заболевания, влекущие за собой изменения ИМТ), а также лиц, страдающих хроническим алкоголизмом, онкологическими заболеваниями, имеющих дисфункцию щитовидной железы, беременных женщин, лиц старше 70 лет.
Всем включенным в исследование проведено клиническое обследование, включающее сбор жалоб и анамнеза, объективный осмотр с измерением антропометрических показателей (масса тела, рост, окружность талии — ОТ и окружность бедер — ОБ) и артериального давления (АД). ИМТ вычисляли по формуле: ИМТ = масса тела (кг)/рост2 (см). Ожирение констатировали при ИМТ ≥30 кг/м2. Метаболический синдром определяли на основании модифицированных критериев АТР III [13]. Все обследованные были также опрошены по опроснику финской диабетической ассоциации для оценки риска развития СД [14], включающего вопросы о потреблении овощей (каждый или не каждый день) и физической активности (более или менее 30 мин в день).
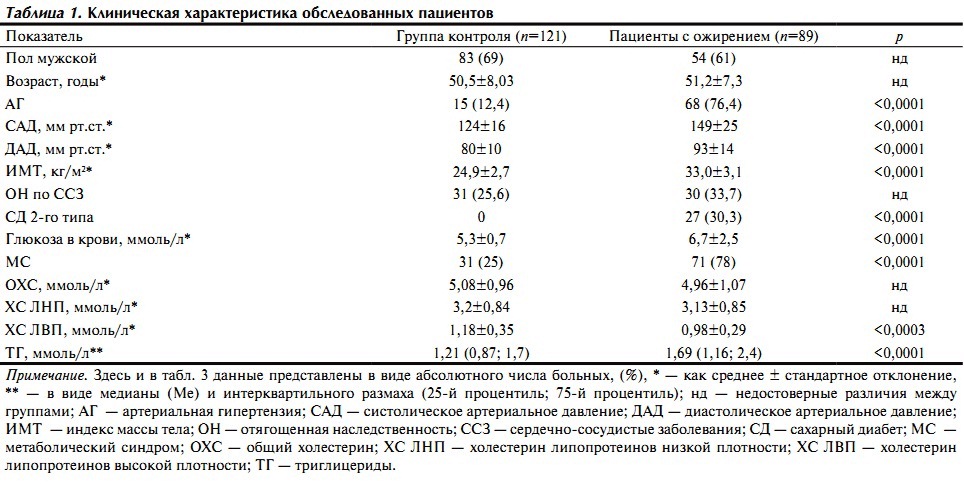
Клиническая характеристика обследованных пациентов представлена в табл. 1.
Таблица 1. Клиническая характеристика обследованных пациентов
По полу и возрасту статистически значимых различий между группами не отмечалось. Согласно критериям деления пациентов по группам у лиц с ожирением имелся более высокий ИМТ, чаще встречались метаболический синдром, АГ с более высокими систолическим и диастолическим АД, СД 2-го типа и повышение уровня глюкозы в крови натощак, а также более высокие уровни три-глицеридов (ТГ) и снижение концентрации холестерина (ХС) липопротеидов высокой плотности (ЛВП).
Биохимические показатели определяли на автоанализа-торе Sinhron СХ4-DELTA («Beckman», США). Кровь для определения биохимических показателей и генетического исследования брали утром натощак после 12-часового голодания. В полученных образцах определяли уровни глюкозы и липидов (общий ХС — ОХС, ХС ЛВП, ТГ). Уровень ХС липопротеинов низкой плотности (ЛНП) определяли по формуле Фридвальда [15].
Таблица 2. Распределение генотипов и аллелей полиморфизма С825Т гена субъединицы β3 белка G у обследованных лиц
Выделение ДНК и генетический анализ. ДНК выделяли из клеток крови с использованием набора для экстракции ДНК Nucleon BACC3 («Amersham Pharmacia Biotech», Швеция). Полиморфизм С825Т гена субъединицы β3 белка G определяли с помощью полимеразной цепной реакции на амплификаторе Hybaid с использованием специфических праймеров (прямой 5'TGA CCC ACT TGC CAC CCG TGC 3', обратный 5'GCA GCA GCC AGG GCT GGC 3) и последующей рестрикцией продуктов полимеразной цепной реакции ферментом BseDI. В результате рестрикции получены следующие фрагменты: TT — 268 п. н., TC — 268+152+116 п. н. и CC — 152+116 п.н. Сканирование рестрикционных фрагментов в 3% агаровом геле и анализ полученных результатов осуществляли на имидж-денситометре GelDoc-It («UVP», США).
Статистический анализ проводили с помощью пакета программ SPSS 17.0. Сравнение переменных с нормальным распределением выполняли при помощи t-критерия Стьюдента, данные представлены как среднее ± стандартное отклонение. Переменные с непараметрическим распределением сравнивали при помощи критерия Манна—Уитни, данные представлены в виде медианы и интерквартильного размаха (25-й процентиль; 75-й процентиль). Для выявления взаимосвязи качественных переменных применяли критерий χ2 и высчитывали отношение шансов (ОШ) с расчетом 95% доверительного интервала (ДИ). Для выявления основных факторов, влияющих на развитие ожирения в исследуемой группе, применяли логистическую регрессионную модель с пошаговым включением переменных. Критерием статистической значимости считались р<0,05.
Результаты
Частота генотипов и аллелей полиморфизма С825Т гена субъединицы β3 белка G у обследованных лиц представлена в табл. 2.
Распределение генотипов и аллелей находилось в равновесии Харди—Вейнберга. Среди обследованных лиц наиболее часто выявлялся гетерозиготный генотип СТ.
Гомозиготный генотип ТТ встречался относительно редко (менее 5%), в связи с чем для проведения статистического анализа лица с наличием мутантного аллеля были объединены в одну группу.
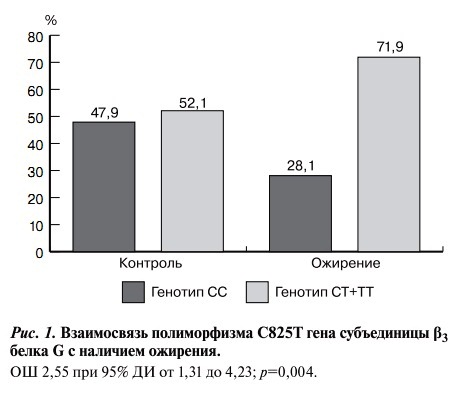
У пациентов с ожирением наличие генотипов СТ+ТТ встречалось достоверно чаще, чем у лиц без ожирения. Носительство мутантного аллеля увеличивало риск развития ожирения более чем в 2,5 раза (рис. 1).
Рис. 1. Взаимосвязь полиморфизма С825Т гена субъединицы β3 белка G с наличием ожирения
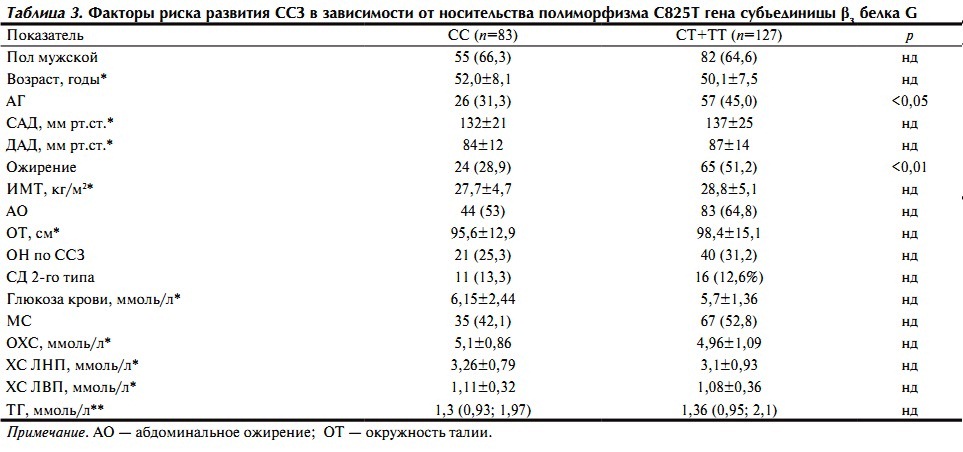
Для выявления взаимосвязи изучаемого полиморфизма с другими факторами риска развития сердечно-сосудистых заболеваний (ССЗ) все пациенты были разделены на 2 группы в зависимости от наличия или отсутствия аллеля 825Т (табл. 3).
Таблица 3. Факторы риска развития ССЗ в зависимости от носительства полиморфизма С825Т гена субъединицы β3 белка G
Отмечено, что у носителей аллеля Т достоверно чаще встречалась АГ. Тем не менее по уровню АД лица обеих групп были сопоставимы. Ожирение также чаще выявлялось у лиц с аллелем 825Т; при этом такие показатели, как ИМТ и ОТ, между группами существенно не различались.
Абдоминальное ожирение по критериям АТР III встречалось несколько чаще у носителей аллеля Т, однако различия не достигали уровня статистической значимости. По другим факторам риска развития ССЗ, включая показатели липидного и углеводного обмена, существенных различий также не выявлено.
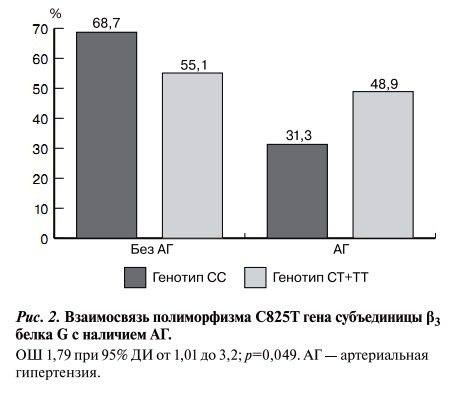
При оценке взаимосвязи между изучаемым полиморфизмом и АГ выявлено, что наличие аллеля Т увеличивает риск обнаружения АГ в 1,79 раза (рис. 2).
Рис. 2. Взаимосвязь полиморфизма С825Т гена субъединицы β3 белка G с наличием АГ
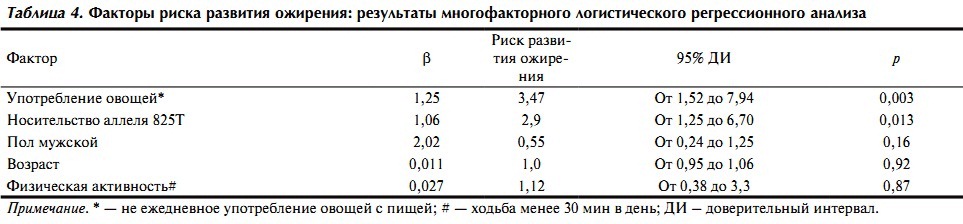
Для оценки влияния различных факторов на развитие ожирения был проведен логистический регрессионный анализ с включением в качестве зависимой переменной ожирения, а в качестве независимых переменных — факторов, способных влиять на его развитие (пол, возраст, употребление овощей: каждый день (0), не каждый день (1), физическая активность (более (0) или менее 30 мин в день (1)) и носительство аллеля 825Т).
По результатам анализа из включенных переменных отсутствие ежедневного употребления овощей повышало риск развития ожирения почти в 3,5 раза, а наличие алле-ля 825Т — в 2,9 раза независимо от пола, возраста и уровня физической активности (см. табл. 4). При построении аналогичной регрессионной модели для АГ аллель 825Т терял свою статистическую значимость после введения в уравнение в качестве независимой переменной ожирения, что косвенно может указывать на опосредованное влияние аллеля 825Т на повышение АД через развитие ожирения.
Таблица 4. Факторы риска развития ожирения: результаты многофакторного логистического регрессионного анализа
Обсуждение
Белок G играет важную роль в обменных процессах, регулируя передачу сигналов от различных рецепторов с поверхности клетки. В неактивном состоянии белок G состоит из 3 субъединиц (α, β и γ) и через субъединицу α связан с гуанозинтрифосфатом (ГТФ). При активации белка происходит переход ГТФ в гуанозиндифосфат (ГДФ) и отделение субъединицы α от комплекса β—γ. В зависимости от типа субъединицы α происходит активация (αs) или ингибирование (αi) различных внутриклеточных молекул, запуская каскад сигналов трансдукции [3]. Среди основных процессов, на которые воздействует белок G, — активация/ингибирование аденилатциклазы, различных изоформ фосфолипазы, регуляция трансмембранного потока ионов К+, Nа+, Са2+, Н+ и каскада митоген-активирующих протеинкиназ и т.д. [4-5]. Экспрессия субъединицы β3 белка G отмечается во всех тканях организма и является ключевым компонентом в передаче сигналов трансдукции. Ген, кодирующий субъединицу β3, локализован в 12-й хромосоме. Замена цитозина тимидином в 825-й позиции не влияет на аминокислотную последовательность в белке, однако приводит к синтезу укороченной субъединицы β3, что вызывает усиление сигналов внутриклеточной трансдукции, вероятнее всего, в результате образования более тесной связи β3—αi [6].
В обследованной нами группе этнических киргизов превалировал гетерозиготный генотип СТ, гомозиготный мутантный генотип встречался относительно редко, и частота аллеля 825Т составила 32,4%. По данным литературы, частота аллеля 825Т сильно варьирует в зависимости от этнической и расовой принадлежности. Так, чаще всего гомозиготный генотип 825ТТ обнаруживался на африканском континенте, где частота мутантного аллеля варьирует от 74 до 91% [7]. В азиатских популяциях частота аллеля составляет в среднем 46% (от 42 до 55%), у лиц европейского происхождения — 33% (21—38%). В странах постсоветского пространства данные по изучаемому полиморфизму доступны для России, где преобладал генотип СС, а частота аллеля Т составила 21%, и для Узбекистана, где частота генотипов СС и СТ была примерно одинаковой, распространенность аллеля Т составила 36% [7].
В нашем исследовании в киргизской этнической группе обнаружена взаимосвязь между носительством аллеля 825Т субъединицы β3 белка G и ожирением. Данная ассоциация также выявлена в большинстве других исследований. Так, в исследовании W. Siffert и соавт. [7] показано, что ОШ выявления избыточной массы тела у носителей генотипа ТТ в среднем колеблется от 2 до 3, причем данная ассоциация отмечалась во всех 3 этнических популяциях, включенных в исследование: у европейцев (ОШ 2,5 при 95% ДИ от 1,1 до 6,1), у азиатов в китайской популяции (ОШ 1,8 при 95% ДИ от 1,0 до 3,1) и у африканского населения (ОШ 2,7 при 95% ДИ от 1,4 до 5,3). Кроме того, было показано, что в африканской популяции данная ассоциация была более выраженной у городских жителей по сравнению с сельскими, что указывает на усиление влияния факторов окружающей среды у лиц с генетической предрасположенностью. При изучении фармакогенетических особенностей отмечено более выраженное снижение массы тела у лиц с генотипами СТ и ТТ при проведении немедикаментозных вмешательств (гипокалорийная диета, увеличение физической активности) [16, 17], а в некоторых из них этот эффект усиливался при добавлении терапии сибутрамином [17, 18].
Возможными механизмами развития ожирения у носителей аллеля 825Т могут быть снижение липолитической активности жировых клеток в ответ на катехоламины [19, 20] и увеличение адипогенеза через избыточную стимуляцию через pertussis toxin-sensitive (РТХ)-рецепторы [21, 22]. Учитывая высокую распространенность аллеля 825Т у лиц азиатского происхождения и на африканском континенте, полиморфизм С825Т гена субъединицы β3 белка G считают «энергосберегающим» полиморфизмом, и его влияние на развитие ожирения согласуется с теорией J. Neel [23], согласно которой гены, исторически отвечающие за накопление жировой ткани в организме, в современных условиях способствуют развитию ожирения, инсулинорезистентности и СД 2-го типа. В Кыргызстане распространенность ожирения растет, что также может быть связано с изменением образа жизни. Исторически киргизские племена были кочевниками, их рацион всегда был богат жирами животного происхождения, что было необходимо для совершения переходов. В настоящее время традиции питания существенно не изменились, однако физическая активность населения снизилась. Это привело к существенному росту распространенности ожирения среди коренного населения, что, возможно, связано с накоплением неизрасходованной энергии и отсутствием генетической защиты.
В нашем исследовании в группе лиц с генотипами СТ+ТТ также чаще выявлялась АГ. Однако при проведении логистического регрессионного анализа статистически значимое влияние аллеля 825Т терялось в случае добавления в регрессионное уравнение в качестве независимого фактора ожирения. Это позволяет сделать предположение об опосредованном влиянии изучаемого полиморфизма на развитие АГ через развитие ожирения. Взаимосвязь между ожирением и АГ подтверждена многочисленными исследованиями [1]. Результаты исследований по изучению взаимосвязи между полиморфизмом С825Т и АГ противоречивы. Некоторые из них подтверждают ассоциацию аллеля 825Т с развитием АГ, гипертрофии левого желудочка [8, 10, 24-26] и атеросклероза [27], в то время как другие — нет [28-32]. В основном положительная взаимосвязь с аллелем Т выявлена в европейских популяциях, в то время как у лиц азиатского происхождения данная взаимосвязь отмечается значительно реже. Следует отметить, что в узбекской популяции, наоборот, носительство генотипа СС ассоциировалось с наличием АГ, и в контрольной группе пациентов с нормальным АД генотип СС вообще не встречался [33]. Мета-анализ 34 исследований с включением 14 094 пациентов с АГ и 17 760 контрольных лиц показал увеличение риска развития АГ у носителей генотипов СТ+ТТ в 1,17 раза (95% ДИ от 1,06 до 1,29), причем данная взаимосвязь была существенно выше после исключения из анализа исследований, проведенных на азиатских популяциях [9].
Возможные механизмы развития АГ у лиц с полиморфизмом С825Т изучены не до конца. Предполагается, что влияние данного полиморфизма на повышение АД осуществляется через механизмы «замедленного» действия, возможно, как результат гиперактивации РТХ-рецепторов и нарушения регуляция трансмембранного потока ионов Nа+, H+ и Са2+ [34]. Показано также, что аллель 825Т субъединицы β3 белка G ассоциировался с низкой концентрацией ренина в крови [35]. Как известно, пациенты с АГ и сниженным уровнем ренина характеризуются повышенной чувствительностью к потреблению поваренной соли с пищей и задержкой жидкости в организме. Интересно, что при ожирении также отмечается задержка поваренной соли и жидкости в организме. Результаты фармакогенетических исследований показали наличие более выраженного гипотензивного эффекта при применении тиазидных диуретиков у носителей аллеля 825Т [36, 37], в то время как существенных различий в ответ на терапию β-адреноблокаторами у носителей генотипов СС, СТ и ТТ не выявлено [38, 39].
Таким образом, в проведенном нами исследовании в группе этнических киргизов носительство аллеля 825Т субъединицы гена β3 белка G ассоциировалось с наличием ожирения.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}