
Изучение механизмов естественной защиты миокарда не теряет своей актуальности до настоящего времени.
Известно, что кратковременные эпизоды ишемии— реперфузии существенно уменьшают величину инфаркта миокарда, возникающего в результате последующей продолжительной ишемии [1]. Одним из многочисленных триггеров ишемического прекондиционирования является норадреналин (НА). Участие его в прекондиционировании было продемонстрировано в экспериментах с предварительным введением кроликам резерпина (для истощения запасов НА в пресинаптических нервных терминалях). Только в группе с ишемическим прекондиционированием без применения резерпина обнаружено уменьшение зоны инфаркта миокарда. У животных, получавших резерпин, величина зоны инфаркта при использовании прекондиционирования и без него не различалась. На основании этого авторы сделали вывод, что освобождение НА играет важную роль в защите миокарда, обеспечиваемую прекондиционированием [2]. В то же время некоторые исследователи не отметили влияния истощения катехоламиновых депо на эффект прекондиционирования [3]. Интересным свойством НА является его способность выступать в качестве как пускового механизма естественной защиты, так и фактора, повреждающего миокард.
Целью настоящего исследования было изучение особенностей метаболизма НА миокарда при длительных ишемии и реперфузии после ишемического преконди- ционирования у крыс в остром эксперименте.
Материал и методы
Исследование выполнено на 32 крысах-самцах Вистар со средней массой тела 393,5±10,6 г, содержавшихся в обычных условиях вивария. Эксперименты проводили в соответствии с «Правилами проведения работ с использованием экспериментальных животных» (Приложение к приказу Министерства здравоохранения СССР от 12.08.1977 г. № 755). Детально методики описаны ранее [4]. Ниже следует краткое описание методов.
Микродиализ.В экспериментах использовали линейные микродиализные зонды на базе полиакрилнитриловой мембраны (CGH Medical Inc., Lakewood, США) длиной 6 мм с внешним диаметром около 300 мкм и с критической массой пропускания 29000 Да. С обеих сторон к мембране приклеивали входной и выходной полиамидные капилляры с внутренним диаметром около 150 мкм (Microlumen, США). Для уменьшения «мертвого объема» и обеспечения прочности и несминаемости мембраны внутрь зонда вставляли нейлоновое волокно диаметром 70 мкм. Подачу перфузата осуществляли насосом MD-1001 (BAS, США) со скоростью 2 мкл/мин. Пробы собирали 1 раз в 10 мин.
Аналитическая процедура. НА диализата определяли при помощи жидкостной хроматографии высокого разрешения с электрохимической детекцией. Система состояла из насоса Shimadzu LC-10ADvp, инжектора Rheodyne 9125 с петлей 10 мкл и колонки Hypersil (C18, 2x150 мм, 5 мкм). Скорость подачи элюента составляла 0,07—0,1 мл/мин. Электрохимическое детектирование осуществляли сдвоенным электродом со стеклоуглеродными пластинами диаметром 3 мм (CC-5) и контроллерами LC-4B (BAS, США). Потенциалы верхнего и нижнего по течению элюента электродов устанавливали соответственно +0,6 и +0,02В относительно референтного электрода Ag/AgCl. Хроматограммы накапливали и обрабатывали при помощи системы Мультихром (Амперсенд, Москва). Анализ концентрации НА в некоторых, а дигидроксифенилгликоля — в большинстве экспериментов выполняли по показаниям второго, более устойчивого к загрязнениям, электрода.
Подвижная фаза состояла из 0,05М одноосновного фосфата натрия, 0,05М лимонной кислоты (Fisher Scientific, США), 80 мг ЭДТА (Aldrich, США) и 500 мг октилсульфоната натрия (Sigma, США) на 1 л при рН 5,6, устанавливаемом при помощи 6M NaOH. Подвижную фазу фильтровали через нейлоновые фильтры 0,2 мкм и дегазировали вакуумом. Метанол, дополнительно дистиллированный, добавляли до концентрации 12% (v/v). Сквозная чувствительность системы была на уровне 0,5 пкг НА в 10 мкл образце (отношение сигнал/шум 3:1).
Хирургическая подготовка и вживление микродиализных зондов. Животным под уретановым наркозом (2,0±0,07 мг/кг внутрибрюшинно) производили трахеотомию. Искусственную вентиляцию легких осуществляли при помощи аппарата Model 683 (Harvard Apparatus, США) смесью воздуха с 40% кислорода. В течение эксперимента контролировали электрокардиограмму. Температуру тела крысы, измеряемую в прямой кишке, поддерживали путем подогрева операционного стола.
При вживлении микродиализного зонда левостороннюю торакотомию выполняли в пятом межреберье. После рассечения перикарда с помощью нейлоновой нити формировали петлю вокруг левой нисходящей коронарной артерии на расстоянии 1—2 мм ниже ушка левого предсердия. Оба конца петли вводили в отверстия пластикового окклюдера. Диализный зонд вживляли в зону ишемии, которая обнаруживалась по обесцвечиванию ткани миокарда при кратковременном затягивании нитей окклюдера. Микродиализный зонд протягивали через зону ишемии миокарда при помощи инъекционной иглы диаметром 0,4 мм, подклеенной к началу выходного капилляра. После вживления зонда на выходной капилляр до упора в миокард надевали силиконовую трубку диаметром 0,6 мм, которая препятствовала подвижкам зонда в миокарде. Сбор образцов диализата для определения НА и дигидроксифенилгликоля осуществляли каждые 10 мин.
Протокол эксперимента. Животные были случайным образом разделены на 2 группы. В экспериментальной группе выполняли ишемическое прекондиционирование — 2 эпизода: окклюзия (5 мин) — реперфузия (10 мин). После этого осуществляли продолжительную (60 мин) окклюзию и реперфузию (60 мин). В контрольной группе длительной окклюзии предшествовал период покоя, по длительности соответствовавший периоду прекондиционирования в 1-й группе (30 мин). Затем следовали окклюзия (60 мин) и реперфузия (60 мин).
Через 30 мин после имплантации микродиализного зонда в миокард в течение 30 мин осуществляли сбор образцов диализата для определения НА и его метаболита дигидроксифенилгликоля (3 раза по 10 мин). Ранее нами было показано [5], что абсолютные значения концентраций НА на этапах исследования в значительной степени зависят от локализации микродиализного зонда, определяющей концентрацию НА в первой пробе. Это дало нам основание нормализовать результаты измерения концентрации веществ и выразить их в процентах от первой пробы, полученной к 40-й минуте после имплантации микродиализного зонда. Стабилизация уровней НА и метаболита устанавливалась к 60-й минуте после имплантации. Поэтому в качестве исходного значения рассматривали величину отношения показателей третьей пробы (60-я минута) к показателям первой (40-я минута после имплантации зонда).
При анализе данных использовали критерий Вилкоксона для парных сравнений или критерий Манна—Уитни с уровнем достоверности 0,05. Результаты представляли в виде М±m (среднее ± ошибка среднего) при помощи программ Statistica for Windows.
Результаты и обсуждение
Концентрация НА при первом измерении в экспериментальной группе составила 0,16±0,016 нг/мл, в контрольной группе — 0,13±0,016 нг/мл. Концентрация дигидроксифенилгликоля составила при первом измерении соответственно 0,21±0,036 и 0,19±0,032 нг/мл. Средние исходные концентрации НА в диализате не различались у животных контрольной и экспериментальной групп, составляя 22,4±5,1 и 19,8±4,3% соответственно.
Ишемическое прекондиционирование не приводило к существенному изменению концентрации НА в диализате. На этом этапе исследования не было отмечено достоверных различий между группами (рис. 1). По данным других исследователей, также не наблюдалось увеличения уровня НА в интерстициальной ткани миокарда при коротких циклах прекондиционирования [6]. Лишь при увеличении продолжительности кондиционирующей ишемии до 10 мин отмечено некоторое нарастание концентрации НА [7].
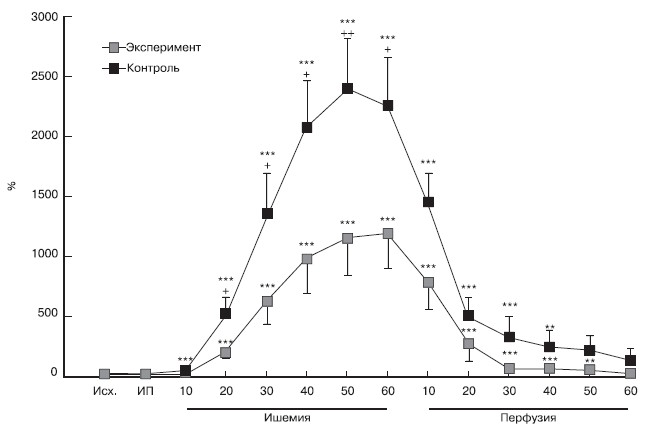
Рисунок 1. Динамика концентраций НА в интерстициальной ткани миокарда во время ишемии и реперфузии.
1 — первое измерение, к которому нормировали концентрации норадреналина на этапах исследования (40-я минута эксперимента); Исх.— исходные показатели — перед ишемическим прекондиционированием; ИП — показатели после ишемического прекондиционирования; НА — норадреналин. Здесь и на рис. 2 различия достоверны по сравнению с экспериментальной группой (+ — p<0,05; ++ — p<0,02), по сравнению с исходным уровнем (* — p<0,05; ** — p<0,02; *** — p<0,01).
Во время продолжительной ишемии отмечено постепенное нарастание концентрации НА в диализате. Такая динамика отмечена как в экспериментальной, так и в контрольной группе, однако в контрольной темп нарастание концентрации НА было существенно выше. Достоверное по сравнению с исходной повышение концентрации НА в диализате отмечено в экспериментальной группе на 20-й минуте ишемии, тогда как в контрольной группе — на 10-й минуте. Уже к 20-й минуте окклюзии прирост концентрации НА в диализате животных контрольной группы достоверно превышал таковой у животных экспериментальной группы (p<0,05).
Достоверное различие между группами сохранялось вплоть до 60-й минуты окклюзии, составляя 1193,0+289,1% в экспериментальной группе и 2262,6+395,0% в контрольной (p<0,05). Во время реперфузии концентрация НА в диализате быстро снижалась в обеих группах. Тем не менее в экспериментальной группе темп снижения был более высоким.
Концентрация НА в интерстициальной ткани миокарда в конце (на 60-й минуте) реперфузии снижалась в экспериментальной группе в 37 раз (p<0,05) по сравнению с концентрацией НА на 60-й минуте окклюзии, тогда как в контрольной группе — лишь в 15,8 раза (p<0,05).
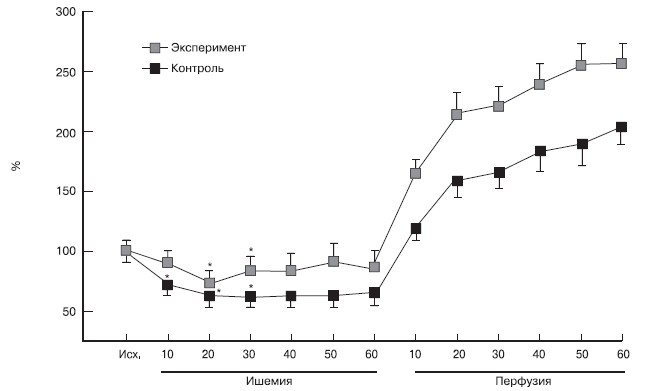
С началом ишемии концентрация дигидроксифенилгликоля в диализате уменьшалась в обеих обследованных группах (рис. 2). В контрольной группе снижение происходило быстрее — достоверные различия по сравнению с исходным значением отмечены уже к 10-й минуте окклюзии (p=0,0086). Сниженная концентрация дигидроксифенилгликоля сохранялась в этой группе вплоть до окончания ишемии. В экспериментальной группе динамика концентрации дигидроксифенилгликоля несколько отличалась. Понижение ее при ишемии происходило на 20 мин позже, чем в контрольной группе (p=0,044). Тем не менее на последующих этапах окклюзии концентрация дигидроксифенилгликоля в диализате несколько увеличивалась и уже не отличалась от исходной до окончания локальной ишемии.
Рисунок 2.Динамика концентраций дигидроксифенилгликоля в интерстициальной ткани миокарда во время ишемии и реперфузии.
Исх.— исходные показатели — перед ишемическим прекондиционированием.
О том, что НА играет существенную роль в ишемическом повреждении миокарда, известно уже давно [8]. Было показано, что концентрация интерстициального НА при ишемии многократно возрастает [9—11]. В эксперименте на изолированных папиллярных мышцах отмечено, что освобождение эндогенных катехоламинов при ишемии ухудшает их сократительную способность, усиливает оглушение и препятствует эффектам клинических доз β-адреноблокаторов, что снижает их защитное противоишемическое действие [12].
При изучении токсических эффектов экзогенного НА на изолированных сердцах кроликов обнаружено, что НА влияет на величину инфаркта миокарда дозозависимо. При низких концентрациях (10-7 моль/л) размер инфаркта не увеличивался. При более высоких (10-6 моль/л, 10-5 моль/л) отмечалось увеличение зоны инфаркта [13].
В качестве одного из объяснений цитотоксичности НА была представлена положительная корреляция уровня медиатора с образованием свободных гидроксильных радикалов в интерстициальной ткани миокарда [14]. Существует мнение, что выработка свободных радикалов зависит не столько от самого НА, сколько от его метаболита, образующегося в результате действия моноаминоксидазы А — 3,4-дигидроксифенилгликольальдегида. Именно в реакциях с этим субстратом образуются свободные радикалы при оксидативном стрессе [15].
В то же время оказалось, что введение экзогенного НА воспроизводит защитный эффект ишемического прекондиционирования, который сохраняется и после прекращения гемодинамических эффектов НА [16]. Известно, что в физиологических условиях НА выводится из синаптической щели преимущественно посредством механизма обратного захвата с помощью белка-переносчика. Обратный захват НА играет существенную роль и в симпатической нейротрансмиссии во время ишемии миокарда, когда в начале ишемии отмечается увеличение элиминации НА вследствие активации обратного захвата. Увеличиваются как функциональная активность этого механизма, так и плотность белка-переносчика обратного захвата [17]. Тем не менее после 10-минутной ишемии концентрация НА в интерстициальной ткани миокарда начинает нарастать.
Высказано предположение, что повышение концентрации НА в интерстициальной ткани миокарда при ишемии связано с реверсией его обратного захвата и зависит от двух условий: 1) увеличения концентрации этого медиатора в цитоплазме симпатического нейрона и 2) интранейронального накопления натрия. Сочетание этих условий необходимо для транспорта НА через плазматическую мембрану с включением в процесс транспортера обратного захвата с реверсией нормального направления транспорта [9].
Полученные нами данные свидетельствуют о существенной задержке накопления НА при длительной ишемии в прекондиционированном миокарде. Непосредственной причиной этого может быть сохранение у такого миокарда нормальной деятельности механизма обратного захвата в течение продолжительного времени. Без прекондиционирования нарушение трансмембранного транспорта НА с использованием переносчика у крыс наступает уже после 50-минутной ишемии [4, 9]. Наши эксперименты продемонстрировали существование различий между прекондиционированным и контрольным миокардом до конца 60-минутного периода. Кроме того, о сохранности механизма трансмембранного транспорта свидетельствует и более резкое снижение концентрации интерстициального НА во время реперфузии у прекондиционированных крыс.
Не меньший, чем динамика интерстициального НА, интерес представляют его изменения внутри нервных окончаний. С целью изучения таких изменений традиционно используется измерение концентрации дигидроксифенилгликоля в интерстициальной ткани миокарда [18]. В симпатических нервных окончаниях свободный НА аксоплазмы под воздействием моноаминоксидазы превращается исключительно в дигидроксифенилгликоль. В силу липофильности этот метаболит легко проникает через мембрану симпатических нервных окончаний вследствие простой диффузии [18]. Таким образом, концентрация дигидроксифенилгликоля в интерстициальной ткани миокарда будет отражать динамику НА внутри аксона.
Во время ишемии отмечалось снижение концентрации дигидроксифенилгликоля в диализате, которое было обусловлено, вероятно, снижением активности моноаминоксидазы, являющейся кислородзависимым ферментом [19]. Тем не менее, хотя концентрация дигидроксифенилгликоля в диализате и уменьшалась, она составляла не менее 60—70% от исходного уровня, что свидетельствует о сохраняющемся, хотя и пониженном дезаминировании НА. Нельзя исключить вероятность того, что поддержание метаболизма связано с остаточным кровотоком в ишемическом регионе, который, по некоторым данным, может достигать 20% от исходного [20].
Тенденция к более высокой концентрации дигидроксифенилгликоля в экспериментальной группе может быть обусловлена сохраняющимся после прекондиционирования нормальным функционированием механизма обратного захвата и, соответственно, большим количеством свободного НА в аксоплазме, подвергающегося дезаминированию [18, 21].
Сразу после начала реперфузии в обеих группах отмечено повышение концентрации дигидроксифенилгликоля в диализате (p<0,05 по сравнению с исходной). Это нарастание было более выражено в экспериментальной группе. По сравнению с предыдущим этапом прирост концентрации дигидроксифенилгликоля составил на 10-й минуте 92%, тогда как в контрольной — 86% (p<0,05). Полученные результаты свидетельствуют о том, что активность моноаминоксидазы даже после продолжительной ишемии во время реперфузии быстро восстанавливается.
Более высокий уровень дигидроксифенилгликоля в реперфузионном периоде в экспериментальной группе по сравнению с контролем обусловлен, вероятно, повышенным количеством свободного НА в аксоплазме вследствие более эффективного функционирования механизма обратного захвата во время ишемии.
Таким образом, двукратное ишемическое прекондиционирование эффективно тормозит накопление НА в интерстициальной ткани миокарда при длительной ишемии. Вероятно, после ишемического прекондиционирования нормальный механизм обратного захвата НА функционирует более продолжительное время, однако депонирование НА в везикулы нарушается, поэтому значительная его часть остается в свободном состоянии в аксоплазме, где и подвергается дезаминированию с участием моноаминоксидазы.



{kind=link}
{kind=link}